
Accueil > 02 - Livre Deux : SCIENCES > Développement et génétique > Une des plus grandes révolutions du Vivant : l’émergence de la cellule eucaryote
Une des plus grandes révolutions du Vivant : l’émergence de la cellule eucaryote
mercredi 7 août 2019, par
Stephen Jay Gould dans « Comme les huit doigts de la main » (chapitre « Les grandes modalités de l’évolution ») :
« De nombreux biologistes situent la grande division au sein de la nature, non pas entre les plantes et les animaux, ni même entre les organismes unicellulaires et multicellulaires, mais, au sein même des unicellulaires, entre les procaryotes et les eucaryotes. »
« Sciences de la Terre et de l’Univers », ouvrage collectif dirigé par Jean-Yves Daniel et André Brahic :
« Mais comment passe-t-on d’une cellule simple, au noyau nu et diffus, à une cellule eucaryotique compartimentée, à vrai noyau et organites cellulaires ? »
Une des plus grandes révolutions du Vivant : l’émergence de la cellule eucaryote
Il est certain que l’apparition de la photosynthèse, de la sexualité, de la cellule à noyau et des organismes pluricellulaires sont les principales révolutions du vivant. Toutes ces « apparitions » ont été émergentes, ce qui signifie qu’elles sont de véritables révolutions, qui renversent l’ancien ordre établi, qui fondent sur des bases toutes nouvelles un ordre qui n’est pas un simple bricolage sur le précédent. Cependant, l’émergence d’un ordre nouveau n’est en rien un phénomène incompréhensible pour la science ni un butin pour les mystiques de toutes sortes. L’apparition des cellules eucaryotes n’est nullement irrationnelle même si elle est extraordinairement novatrice.
La vie n’est pas née en une fois mais par toute une série de révolutions qui ne se sont pas déroulées régulièrement mais ont été séparées de longues périodes historiques. Dans son ouvrage « Singularités » De Duve imagine les singularités suivantes du vivant :
1- formation des premières molécules (éventuellement dans l’espace) : chimie abiotique produisant des acides aminés, les pyrophosphates et thioesters
2- production de l’ATP et autres molécules porteuses de l’énergie des interactions du vivant
3- formation des bases U, A, G et C, les mononucléotides NMP et leurs dérivés pyrophosphatés les NTP, qui sont les briques du futur ARN (mais aussi de l’ADN qui n’apparaît que beaucoup plus tard semble-t-il) et appariement des bases. Apparition d’un protométabolisme.
4- apparition de l’ARN (acide ribonucléique) que De Duve appelle « événement charnière » car l’ARN est à l’origine des protéines et des métabolismes. Mais cela ne signifie pas une seule naissance car d’emblée apparaissent de multiples ARN.
5- réplication et transformation de l’ARN par lui-même. Formation des ARN auto-catalytique (l’ARNr ribosomial est la première molécule catalytique, avant les enzymes), ARN messager (ARNm) et de l’ARN de transfert (ARNt). Invention de la variation en même temps que la réplication. L’ARN est à la fois porteur de la mémoire génétique, dépositaire réplicable et agent de cette réplication (ce qui ne sera plus vrai avec l’apparition de l’ADN avec lequel la transcription sera complètement dissociée de la réplication). Début du mécanisme de sélection. Allongement des brins d’ARN. Développement des enzymes et du métabolisme.
6- production des protéines par l’interaction entre diverses sortes de molécules ARN (ARNm, ARNt et ARNr), traduction du langage nucléique en langage protéique, que De Duve nomme la « vraie révolution », « l’événement clef par lequel l’information est entrée dans la vie émergente ».
7- formation de la membrane cellulaire par des protéines membranaires
8- naissance de la cellule vivante (son apparition est particulièrement difficile à situer dans le temps) avec apparition de la croissance et de la multiplication par division.
9- apparition du code génétique : langage de transcription (général au vivant utilisant l’ADN) entre les bases de l’ARN (les NTP) couplées par trois (formant un codon) et les acides aminés (un acide correspondant de façon unique à un codon). Formation de la base thymine T par méthilisation de l’uracile U.
10- apparition de l’ADN (acide désoxyribonucléique), la fameuse double hélice et du double processus de réplication (un brin répliqué d’un seul coup et l’autre par segments reconstitués ensuite) puis revérification par des enzymes correcteurs. L’ordre de l’ADN est issu du désordre et des interactions des ARN qui ne disparaissent pas ensuite mais sont intégrés au processus… Les microARN (non codants) servent à réguler l’expression des gènes.
11- naissance de l’être procaryote (cellule sans noyau) puis eucaryote (cellule à noyau par l’intégration par symbiose à un procaryote d’autre procaryotes constituant mitochondries et chloroplastes). Naissent les familles de procaryotes, les bacteria et les archaea, puis les eucarya (ou eucaryotes, parmi lesquels apparaîtront notamment les animaux, les plantes et les éponges).
12- apparition des êtres pluricellulaires.
Petite chronologie
date inconnue : apparition des ARN
date inconnue : apparition des protéines
date inconnue : apparition de l’ADN
3,5 milliards d’années : apparition de la vie cellulaire ; apparition des premiers stromatolithes
3,2 milliards d’années : apparition des premiers acritarches
3 milliards d’années : apparition de la photosynthèse
2,9 milliards d’années : accrétion des continents (première fois que l’on trouve de vastes zones continentales)
2,7 milliards d’années : première cellules eurcaryotes
2,45 à 2,2 milliards d’années : « grand événement » d’apparition de l’oxygène avec, comme conséquence, l’effondrement du méthane et la destruction massive de formes de vie pour lesquelles l’oxygène est un poison
2,2 milliards d’années : première augmentation de l’énergie solaire (plus 10%)
2,1 milliards d’années : apparition des algues rouges
2 milliards d’années : émergence de la sexualité des bactéries ; apparition des pluricellulaires
1,56 milliards d’années : formation de cellules eucaryotes complètes
Les modifications historiques du vivant sont des ruptures sans continuité possible, qu’il s’agisse de l’apparition de la bactérie, de l’apparition de la cellule eucaryote, de l’apparition des êtres pluricellulaires, de l’apparition de la respiration à oxygène, de l’apparition de la sexualité, de l’apparition des vertèbres, du cerveau et de bien d’autres transformations fondamentales de la structure de l’individu.
Une nouvelle émergence va surgir lorsque les cellules eucaryotes vont à leur tour découvrir l’intérêt de s’associer entre elles pour former une nouvelle société de cellules, comme l’avaient fait les bactéries après 2 milliards d’années de tâtonnement pour aboutir à la cellule eucaryote.
Comme précédemment, il fallait que se forment des cellules spécialisées complémentaires pour qu’au lieu d’aller vivre sa vie chacune pour soi, ces cellules vivent en communauté et collaborent pour former une autre forme de vie dont la complexité se situe à un niveau encore supérieur. Pour réussir cet exploit, il fallait des cellules très sophistiquées, eucaryotes certes, mais aussi totipotentes, c’est-à-dire avec un si large champ de potentialités qu’elles puissent chacune se développer différemment selon l’environnement qui lui serait proposé à l’intérieur du nouvel organisme ! Il fallait aussi que se développent des facteurs de cohésion, d’organisation, de synchronisation, de communication tous indispensables pour que soit viable cette forme de vie des milliards de fois plus complexe que la précédente. Ces facteurs vont devenir prépondérants car la reproduction, la survie ne se fait plus au niveau de la cellule mais de l’organisme multicellulaire tout entier. Toutes les cellules ont le même matériel génétique, mais l’organisme se reproduit globalement grâce à ses gamètes. La sélection va jouer sur l’organisme résultant de l’association de cellules, pas sur les cellules elles-mêmes. C’est donc ces propriétés émergentes qui vont maintenant faire la différence et qui seront l’objet de la diversité sélective. Les cellules eucaryotes sont maintenant performantes, adaptables, totipotentes pour certaines. Ce qui va compter, c’est leur organisation, leur synchronisation, la performance de leur association pour s’adapter aux situations nouvelles et pour se multiplier.
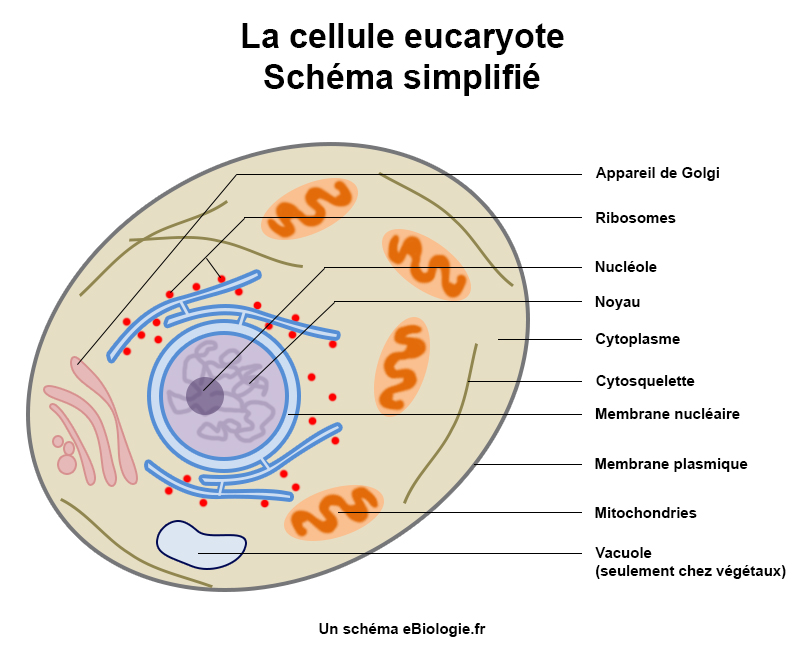
La cellule eucaryote par exemple est faite de composants biochimiques variés, comme les acides nucléiques et les protéines, et est organisée dans des structures limitées comme le noyau de la cellule, diverses organites, une membrane de cellule et le cytosquelette. Ces structures basées sur un flux externe de molécules et d’énergie « produisent » les composants qui, à leur tour, continuent de maintenir la structure contenue.
Le modèle minimal d’un système autopoïétique est une cellule dont la membrane est composé d’un constituant C se dégradant en D et baignant dans un milieu riche en molécules A. Ces molécules A peuvent franchir la membrane et être transformées au sein de la cellule en molécules B pour lesquelles la membrane est imperméable. B peut s’intégrer à la membrane pour se transformer en C. Si le débit d’entrée de A et sa conversion en B sont suffisamment grands devant le coefficient de dégradation de C en D alors la cellule se maintient au cours du temps. Ce modèle montre l’importance de la frontière du système (ici la membrane) si celle-ci disparaît le métabolisme et le système entier s’effondre. Il s’agit d’un cercle vicieux : si B s’échappe, sa concentration diminue de telle sorte que la membrane se dégrade de plus en plus vite et que la perte en B augmente. Le métabolisme et la membrane dépendent l’un de l’autre, la structure ne peut se maintenir sans le flux.
Stephen Jay Gould dans « Darwin et les grandes énigmes de la vie » (chapitre « Les cinq règnes de la vie ») :
« Les premières traces de vie ont été découvertes dans les roches vieilles de trois milliards d’années environ. Jusqu’à un milliard d’années avant notre ère, les fossiles ne révèlent que l’existence d’organismes procaryotes ; pendant deux milliards d’années, les matelas d’algues bleues ont été la forme de vie la plus complexe sur terre.
Les opinions divergent en ce qui concerne la suite.
J. W. Schopf, paléobotaniste de l’université de Los Angeles, croit pouvoir affirmer qu’il a découvert, en Australie, des algues eucaryotes dans les roches vieilles d’un milliard d’années environ.
D’autres soutiennent que les micro-organismes de Schopf sont en réalité le produit de la décomposition de cellules procaryotes. Si ces critiques sont exactes, les premiers organismes eucaryotes sont apparus à la fin du précambrien, juste avant la grande « explosion » du cambrien, il ya 600 millions d’années.
Quoiqu’il en soit, la Terre a été le domaine exclusif des organismes procaryotes pendant une période représentant au moins deux tiers de l’histoire de la vie.
Schpf appelle le précambrien, « l’âge de l’algue bleue ».
Margulis a relancé les discussions, il y a quelques années, en défendant une ancienne théorie. L’idée paraît absurde au premier abord, mais on en vient rapidement à l’examiner avec sérieux, et même à y adhérer.
Selon Margulis, les cellules ont été, à l’origine, des colonies d’organismes procaryotes. Le noyau et la mitochondrie, par exemple, étaient au départ des organismes procaryotes indépendants.
Il arrive que des organismes procaryotes envahissent des cellules eucaryotes et vivent en symbiose avec elles.
Presque toutes les cellules procaryotes ont à peu près la même taille que les organites ; la ressemblance entre le chloroplaste de certaines cellules eucaryotes photosynthétiques et certaines algues bleues est frappante.
Enfin, certaines organites possèdent leurs propres gènes, qui leur permettent de se produire indépendamment, vestige de leur statut d’organisme à part entière…
Chez les organismes eucaryotes, les cellules sexuelles ont la moitié des chromosomes des cellules normales. Quand deux cellules sexuelles se joignent pour produire un descendant, la quantité originelle de matériel génétique est reconstituée. Chez les organismes procaryotes, la reproduction sexuelle est rare et inefficace. Elle se fait au hasard, et ne comporte que le transfert de quelques gènes d’une cellule donneuse à une cellule receveuse.
La reproduction asexuelle produit des copies des cellules parentales, à moins qu’une mutation n’intervienne et ne fasse apparaître une variation mineure. Mais les mutations sont rares et la variabilité des espèces asexuelles est trop limitée pour permettre l’évolution. Pendant deux milliards d’années, les matelas d’algues sont restés des matelas d’algues. Mais la cellule eucaryote a fait du sexe une réalité, et c’est pourquoi, moins d’un milliard d’années plus tard, et il y a des hommes, des cafards, des hippocampes, des hippocampes, des pétunias et des moules…
L’augmentation de la diversité et la multiplicité des transitions est peut-être le reflet d’une progression inexorable en direction de choses supérieures. Mais les éléments rassemblés par la paléontologie démentent cette interprétation. Le progrès dans le développement des organismes n’a pas été continu. Il ya eu, au contraire, de très longues périodes de stabilité et une explosion, qui a créé le système dans son entier. Pendant les deux tiers, voire les cinq sixièmes de l’histoire de la vie, la Terre n’a été habitée que par les monères, et rien ne nous permet de dire qu’il y a eu des organismes procaryotes, supérieurs ou inférieurs. De même, aucune structure nouvelle n’est apparue depuis que l’explosion du cambrien a donné naissance à notre biosphère.
L’apparition du système a occupé environ 10% de l’histoire de la vie, pendant l’explosion du cambrien, il y a environ 600 millions d’années. J’en retiendrai deux événements principaux : la formation de la cellule eucaryote – qui rendait la complexité possible en introduisant la variation génétique par l’intermédiaire d’une reproduction sexuelle efficace – et l’envahissement de l’espace écologique par des organismes multicellulaires.
Le mode de vie était calme auparavant et il est redevenu depuis. Il faut considérer la récente apparition de la conscience comme l’événement le plus important depuis l’explosion du cambrien, ne serait-ce qu’en raison de ses conséquences géologiques et écologiques. Les structures nouvelles ne sont pas forcément à l’origine des événements marquants de l’évolution. Les organises eucaryotes continueront à engendrer nouveauté et diversité aussi longtemps que l’une de leurs dernières productions ne se contrôlera pas suffisamment pour assurer un avenir au monde. »
Stephen Jay Gould dans « Comme les huit doigts de la main » (chapitre « Les grandes modalités de l’évolution ») :
« Pour les évolutionnistes, la découverte de la génétique moléculaire la plus fascinante et la plus inattendue a été réalisée dans les années 1960-1970, quand étude après étude, on s’est aperçu que, chez les organismes multicellulaires, seulement un petit pourcentage du patrimoine génétique total consiste en gènes fonctionnels, présents en un seul exemplaire.
La plus grande partie du matériel génétique semble être du « déchet », tout à fait incapable de fournir les informations nécessaires à l’édification et au fonctionnement de l’organisme.
En outre, de nombreux gènes présentent de multiples copies, et c’est un phénomène dont les raisons restent obscures, car elles ne sont pas liées aux fonctions nécessaires du corps.
Néanmoins, il est devenu rapidement clair aux yeux des évolutionnistes que la redondance (sous la forme de copies multiples), présentée par de nombreux gènes, était peut-être un facteur crucial nécessaire à la complexification.
Supposez que l’ancêtre unicellulaire original commun à tous les organismes complexes n’ait possédé qu’une copie par gène, et que chaque gène ait été responsable d’une fonction vitale. Il ne s’agit pas, en fait, d’une simple supposition ; les plus simples des organismes vivants d’aujourd’hui présentent effectivement de telles caractéristiques, et ils constituent les modèles des ancêtres les plus anciens des organismes multicellulaires.
De tels êtres unicellulaires fonctionnent très bien ; ils sont sans doute façonnés avec précision par la sélection naturelle, laquelle les débarrasse de tout trait superflu.
Mais, à présent, voici une énigme pour l’évolution : ces organismes excitent peut-être notre admiration par leur efficacité, mais comment peuvent-ils changer, dans ce sens crucial consistant à ajouter de nouvelles dimensions à la complexité ?
Chacun de leurs gènes code pour quelque chose de vital ; il ne peut se modifier que dans le sens d’une amélioration au sein de son propre domaine fonctionnel. Ce genre de système génétique n’offre aucune flexibilité, aucun jeu, aucune possibilité d’ajouter quelque chose de vraiment nouveau.
Cette énigme a conduit les évolutionnistes à se rendre compte que les copies de gènes multiples pouvaient jouer un rôle fondamental, en permettant l’évolution de la complexité.
Si les gènes présentent plusieurs copies, mais qu’une seule dessert les besoins fonctionnels de l’organisme, alors les autres copies sont libres d’expérimenter, de varier, et d’acquérir de nouvelles fonctions, grâce à des coups de chance occasionnels.
Tout cela est bel et bon, mais nous voici à présent confrontés à une énigme logique : à moins que nous nous méprenions complètement sur la nature fondamentale de la causalité, il n’est pas possible de soutenir que des copies multiples viennent à l’existence parce qu’elles auront un jour une utilité, c’est-à-dire permettront le processus de complexification, des millions d’années plus tard.
Les copies multiples sont la clé du processus de complexification, mais leur apparition dans l’évolution a dû se faire pour d’autres raisons – c’est le principe « des pneus aux sandales » sous-tendant les changements de fonction capricieux.
Ce cas est particulièrement intéressant, parce que la raison initiale de la duplication (l’équivalent de la récolte de caoutchouc pour faire des pneus) n’a sans doute pas eu, par elle-même, grand-chose à voir avec la sélection naturelle, telle qu’elle est traditionnellement conçue, c’est-à-dire comme lutte entre des organismes pour le succès reproductif.
Le phénomène de duplication résulte peut-être d’un processus de sélection à un niveau inférieur, celui des gènes, processus qui reste invisible au niveau plus élevé local.
Les gènes jouent aussi au jeu de la sélection naturelle dans leur univers propre, et ceux qui développent la capacité de se recopier et de changer de place (les transposons ou « gènes sauteurs ») s’assurent un avantage à ce niveau inférieur, tout comme les organismes s’assurent le succès, dans le monde de Darwin, en laissant davantage de descendants survivants.
En fait, la duplication de gènes est peut-être encouragée par le fait qu’elle ne produit aucun effet au niveau du corps, car, en restant invisible au niveau du processus darwinien local, elle peut accumuler des copies supplémentaires « non nécessaires » sur les chromosomes, dans des conditions telles qu’aucune pression négative de la part de la sélection naturelle ne s’y opposera.
Cependant, ces copies de gènes « redondantes » pourront éventuellement être à la source de la complexification ultérieure.
De nombreux biologistes situent la grande division au sein de la nature, non pas entre les plantes et les animaux, ni même entre les organismes unicellulaires et multicellulaires, mais, au sein même des unicellulaires, entre les procaryotes et les eucaryotes.
Les premiers, qui regroupent les bactéries et les algues cyanophycées, sont structuralement plus simples, et n’ont pas d’organites dans leur cellule – ni noyau, ni mitochondries. Les seconds, plus complexe, ont acquis par évolution cette gamme de structures internes qui, dans presque tous les examens de biologie de fin d’études secondaires, fait l’objet d’une inévitable question : nommez tous les organites de la cellule et définissez leurs fonctions.
On considère que l’accroissement de complexité représenté par le passage des procaryotes aux eucaryotes est fondamental, en partie parce que l’on estime que l’organisation eucaryote est le facteur crucial qui a conditionné l’apparition évolutive ultérieure des organismes multicellulaires.
Pour ne citer qu’un seul argument classique : l’évolution darwinienne productrice de complexité a besoin d’abondantes variations dans les caractéristiques des organismes, ceci afin de fournir le matériau nécessaire à la sélection naturelle ; la plupart de ces variations se forment grâce à la reproduction sexuelle, car celle-ci assure le mélange de deux systèmes génétiques différents chez chaque rejeton ; la reproduction sexuelle nécessite un mécanisme pour réaliser de façon exacte la division du matériel génétique, de telle sorte que chaque parent fournissant 50%, les 100% sont reconstitués chez le rejeton ; la méiose, avec sa division réductionnelle des paires de chromosomes, est l’invention biologique qui a permis la division du patrimoine génétique de chaque parent en deux moitiés égales ; les procaryotes, dépourvus de paires de chromosomes et d’autres organites, ne peuvent réaliser une division génétique exacte.
Nous nous trouvons à présent devant la même énigme que celle rencontrée dans le cas précédent : nous comprenons bien que les organismes multicellulaires avaient besoin de l’apparition évolutive des organites, mais les unicellulaires eucaryotes sont apparus au moins 800 millions d’années avant les animaux multicellulaires - de sorte que le progrès dans la complexification représenté par la vie multicellulaire ne peut avoir été la source de l’apparition des organites.
Il existe une théorie en vogue pour expliquer l’apparition évolutive de certains organites (les mitochondries et les chloroplastes), mais, hélas, pas le noyau - Il n’y a, actuellement, pas de théorie satisfaisante pour celui-ci -. Elle fait appel à un processus de symbiose.
Les mitochondries et les chloroplastes ressemblent de manière étrange à des organismes procaryotes complets (ils possèdent leur propre ADN et ont les mêmes dimensions que les bactéries).
Il est presque certain qu’ils ont dû commencer par être des symbiontes au sein des cellules d’autres espèces bactériennes, et se sont intégrés plus tard à celles-ci de façon plus étroite, ce qui a donné la cellule eucaryote (de telle sorte qu’actuellement chaque cellule de notre corps a le statut évolutif de ces anciennes colonies).
Or, on peut soutenir que les ancêtres des mitochondries ont entamé une vie en symbiose parce qu’ils y ont été poussés par le processus local.
Ces bactéries ont ainsi obtenu une protection ou quelque autre avantage, et n’ont donc pas pénétré dans les cellules eucaryotes primitives afin de permettre à la vie multicellulaire complexe d’apparaître par évolution un milliard d’années plus tard.
La symbiose a été réalisée pour des raisons darwiniennes immédiates ; puis la roue a tourné et le caoutchouc de la symbiose a permis de faire les premiers pas en direction de la vie multicellulaire complexe. »
« Sciences de la Terre et de l’Univers », ouvrage collectif dirigé par Jean-Yves Daniel et André Brahic :
« La vie sur Terre aurait donc émergé il y a au moins 3,85 milliards d’années, moins de cent millions d’années après son refroidissement…
Entre 3 et 1,6 milliards d’années, les données deviennent claires et concernent des Procaryotes, ou Monères (bactéries et cyanophycées). Aussi cette période est-elle parfois nommée « ère des monères »…
Les stromatolites précambirens, présents dès – 3,4 milliards d’années (Zimbabwe) et abondants à partir de – 2,8 milliards d’années (groupe de Fortescue en Australie, Afrique du Sud…), ressemblent en tout point aux formes actuelles une superposition de feuillets sub-millimétriques, qui n’est pas une construction biologique, mais une structure laminaire résultant de l’activité d’accumulation de carbonates par des feutrages de cyanophycées.
Il est probable que ces formes anciennes soient le résultat de l’activité de cyanobactéries, mais rien n’est sûr, et certaines bactéries photosynthétiques actuelles construisent des édifices analogues.
Il faut donc rester prudent quant au rôle de pourvoyeur d’oxygène des stromatolites.
La seule conclusion irréfutable est de dire qu’il existe vers – 3,4 milliards d’années des communautés d’organismes autotrophes.
L’apparition de la cellule eucaryote est datée d’environ 1,4 milliard d’années et sa distinction d’avec la cellule procaryote repose à l’état fossile essentiellement sur un critère de taille : sont considérées comme eucaryotes les cellules dont la taille dépasse 60 micromètres. Les plus anciens documents de ce type sont des acritarches.
Mais comment passe-t-on d’une cellule simple, au noyau nu et diffus, à une cellule eucaryotique compartimentée, à vrai noyau et organites cellulaires ?
La découverte d’ADN dans les mitochondries et dans les chloroplastes suggère une certaine autonomie de ces organites.
L’ancienne théorie endosymbiotique ou micromérisme, formulée dès 1883 par Schimper et remise à l’honneur par Margulis dans les années 1970, paraît actuellement la plus propice à rendre compte de la structure complexe de la cellule eucaryote.
Chez les procaryotes il n’existe presque jamais de compartiments délimités à l’intérieur de la cellule et l’ADN se trouve directement dans le cytoplasme par des membranes analogues à celle entourant la cellule. Leur ADN est situé dans l’un des organites, le noyau.
Alors que certains organites de la cellule eucaryote disparaissent pour se reformer lors de la division cellulaire (le Golgi, le réticulum endoplasmique…) d’autres proviennent toujours d’une division d’organites préexistants.
Il s’agit :
– du noyau,
– des mitochondries, responsables de la respiration cellulaire,
– des plastes des végétaux où se déroule la photosynthèse
Cette particularité fait penser à des bactéries se divisant par bipartition dans le cytoplasme : les mitochondries et les plastes seraient des organismes vivants, installés en symbiose dans la cellule.
Divers arguments peuvent être avancés pour étayer cette hypothèse :
– Les organites des eucaryotes sont limités par une membrane lipidique simple sauf trois d’entre eux : le noyau, entouré d’une enveloppe à deux membranes percée de pores, et les plastes et les mitochondries qui sont séparés du cytoplasme par deux membranes continues.
– Les lipides des membranes entourant les mitochondries existent dans la membrane de certaines bactéries, mais pas ailleurs dans la cellule eucaryote.
– Des fonctions métaboliques comme la photosynthèse s’opèrent de façon quasi identique dans les cyanobactéries et les plastes.
– Le génome des plastes et des mitochondries ressemble à celui des procaryotes : il n’est pas isolé au sein de l’organite et se présente sous la forme de plusieurs copies identiques d’une molécule circulaire.
– La comparaison des gènes des organites avec les gènes homologues des procaryotes montre que les mitochondries descendraient toutes d’une bactérie ancestrale unique.
L’acquisition des mitochondries il y a 1,4 à 2 milliards d’années et l’existence de respiration bactérienne antérieure par les cellules eucaryotes marque l’acquisition de la respiration cellulaire.
Sans mitochondries, la cellule eucaryote ne peut utiliser l’oxygène pour oxyder les glucides et produire son énergie.
Associées aux peroxysomes, elles ont permis à la biosphère d’affronter sa première grande crise : le passage d’un environnement aérobie. L’acquisition plus tardive des plastes, entre -1,2 et -1,4 milliards d’années, permettra à la cellule eucaryote d’utiliser l’énergie lumineuse. »
Des procaryotes aux eucaryotes
Des procaryotes aux eucaryotes
Cellule procaryote et cellule eucaryote
L’origine des cellules eucaryotes
Qu’est-ce que la cellule vivante ?