
Accueil > 02 - Livre Deux : SCIENCES > Formation et filiation de l’homme > L’homme, une espèce révolutionnaire
L’homme, une espèce révolutionnaire
lundi 11 janvier 2016, par
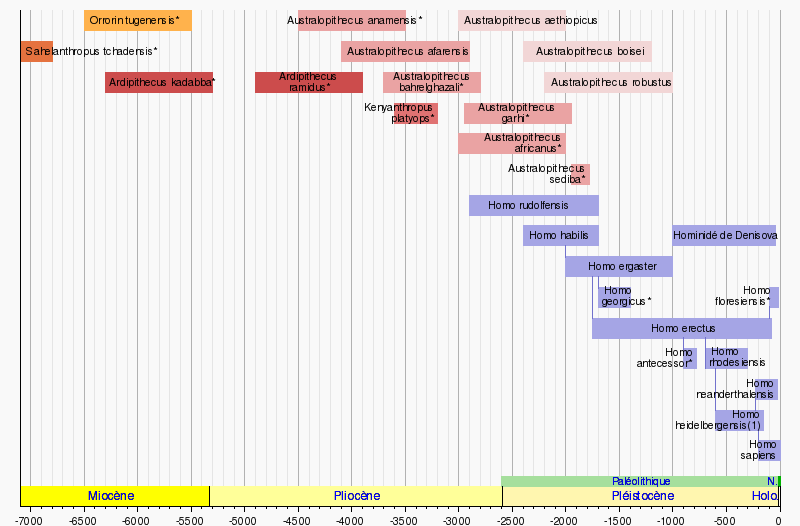
Les multiples révolutions du genre Homo
Il y a eu plusieurs grandes révolutions qui ont mené au genre homo et aussi plusieurs révolutions au sein du genre.
S’il n’existe aujourd’hui qu’une seule espèce du genre homo, l’homme actuel ou homo sapiens sapiens, l’évolution des "homo" a pu permettre de mettre en évidence douze à quinze espèces et il en a existé probablement beaucoup plus. A la plupart des époques, contrairement à aujourd’hui, il a coexisté plusieurs espèces d’homos, et parfois plusieurs sur des sites proches. Par exemple, en Afrique, ont coexisté habilis, rudolfensis et australopitecus boisei. Du vivant de l’homme actuel (homo sapiens sapiens), on n’a rencontré aucun autre homo vivant…
Les homo ont une grande taille corporelle, sont doués de bipédie exclusive, sont adaptés à la vie dans les milieux ouverts.
Le genre homo a notamment été marqué par la fabrication d’outils, même si les grands singes ont des capacités, eux aussi, à en fabriquer et à en utiliser.
Les datations de ces diverses espèces d’homo ne permettent pas de développer un schéma linéaire d’évolution progressive dans lequel une espèce disparaitrait pour en donner une autre. De même que les australopithèques n’ont pas disparu quand les homo apparaissent, ni les autres grands singes, les diverses espèces d’homo ne se succèdent pas.
Il n’est pas juste de présenter la transformation révolutionnaire au sein du genre homo comme une ligne droite habilis-erectus-sapiens.
Il n’est même pas possible de dire qui serait le premier homo, d’habilis ou de rudolfensis. Il n’y a pas nécessairement une seule origine. Les arborescences des révolutions d’espèces ne sont pas à sens unique !
Il est remarquable que la recherche des chaînons manquants de l’évolution ne donne rien de satisfaisant, pour l’homme comme pour la plupart des espèces. En effet, dans les phases évolutives, les divers produits de l’évolution sont d’abord interféconds et on ne peut nullement suivre une évolution linéaire et continue puisque les diverses évolutions échangent encore des gènes.
Les « homo » sont issus d’autres révolutions d’espèces qui ont concerné les « australopitèques » et les « grands singes ».
Deux espèces (rudolfensis et habilis) ont fait la transition entre les homo et les australopithèques. Les considérer ou pas comme des homo est une question pour ceux qui veulent trouver une linéarité là où il n’y en a évidemment pas. Il y a en fait des discontinuités (révolutions) tout au long de l’histoire des humains comme de toute l’histoire des espèces.
Des révolutions donnant un développement inégal et combiné, voilà ce qui semble le schéma pour comprendre le buissonnement de l’évolution.
Bien des preuves génétiques existent de croisements entre espèces homo différentes. Par exemple, les accouplements préhistoriques, il y a environ 40 000 ans, avec des Néandertaliens et des Dénisoviens (nos cousins aujourd’hui éteints qui vivaient en Sibérie), auraient joué un rôle important dans le développement des allergies dont nous souffrons aujourd’hui. La comparaison avec les mêmes séquences d’humains modernes montre que 1 à 4 % des polymorphismes nucléotidiques de l’ADN est commun aux Néandertaliens et aux Homo sapiens eurasiatiques mais pas africains ; ces croisements auraient pu survenir il y a 150 000 à 80 000 ans au Moyen-Orient. Les chercheurs ont trouvé des zones où l’ADN nucléaire des néandertaliens présente des caractéristiques retrouvées chez l’individu de nouvelle guinée, le chinois et le Français mais pas chez les africains. Cela les conduit à penser qu’une partie (faible 4%) du génome des néandertaliens s’est recombinée avec celui d’homo sapiens hors d’Afrique. L’ADN mitochondrial de l’homo de Denisova se situe tout à fait en dehors de la variabilité des homo sapiens et des néerdantaliens. Il présente environ deux fois plus de différences avec Homo sapiens que néandertal et il est également distant des sapiens que des néandertaliens.
• Homo rudolfensis (2,4 millions d’années à 1,9 millions d’années).

• Homo habilis (de 2,45 millions d’années à 1,5 millions d’années).

Les espèces d’homo :
• Homo ergaster (1,9 millions d’années à 1,25 millions d’années).

• Homo georgicus (1,8 million d’années à 1,6 millions d’années).

• Homo erectus : Afrique australe (de 1,8 millions d’années à 300.000 ans).

• Homo cepranensis (800.000 ans).

• Homo antecessor (de 800.000 à 350.000 ans).

• Homo floresiensis (entre 95 000 et 12 000 ans).

• Homo heidelbergensis (entre 600 000 ans et 200 000 ans).

• Homo erectus reilingensis (de 250.000 à 300.000 ans).

• Homo naledi (de 2 à 3 millions d’années).

• L’hominidé de Denisova (entre 1 million d’années et 40 000 ans).

• Homo rhodesiensis (de 300 000 ans à 120.000 ans).

• Homo sapiens, ou « homme de Cro-Magnon » (de 250.000 ans à aujourd’hui).

Parmi lesquels on distingue :
Homo sapiens idaltu (195.000 à 154.000 ans).

Homo neanderthalensis (entre 250.000 et 28.000 ans).


Un lecteur nous fait remarquer qu’il y a une contradiction apparente à parler d’espèces différentes d’homo et à affirmer qu’elles se seraient interfécondées. Et il a raison. Il y a une contradiction mais pas diamétrale : dialectique au sein même de la notion d’espèce. Voir ici : L’espèce vivante, une notion dialectiquement contradictoire
Les paradoxes de la notion d’espèce
Ce qui est faux, c’est de considérer que les espèces seraient séparées par des barrières infranchissables. Voir ici : Les espèces séparées par des barrières infranchissables ?
L’espèce nouvelle n’a pas besoin d’avoir un matériel génétique fondamentalement nouveau pour naître. Sinon, elle ne pourrait pas apparaître, le « nouveau » étant incapable de féconder un seul des anciens. C’est au sein de l’ancienne espèce qu’apparaît la nouveauté et donc avec le matériel génétique de l’ancien. Il suffit qu’il y ait eu une modification d’utilisation de ce matériel génétique qui ait été acceptée et non bloquée par inhibition, par exemple du fait d’un stress environnemental inhibant les protections internes hostiles à la nouveauté. Voir ici : Le fonctionnement révolutionnaire du vivant
La nouveauté peut être possible au sein de l’ancien matériel génétique qui est plus contradictoire qu’il n’y paraît et avoir été bloquée par des mécanismes de protection. Dès que cette protection cesse, la nouveauté peut apparaître au sein même de l’ancien capital génétique. Il y a alors saut brutal : une révolution. Voir ici : Evolution ou révolution des espèces ?
Le changement brutal d’espèces
L’espèce est une structure émergente issue des contradictions précédentes et de leur résolution révolutionnaire.
Les différentes révolutions du passé des origines de l’espèce sont ainsi intégrées dans son développement. Voir ici : L’espèce humaine, comme produit du développement inégal et combiné de la vie animale
La nouvelle espèce n’apparaît pas à cause d’un matériel génétique déjà complètement différent. Il va se différencier de plus en plus s’il n’y a pas de fécondation croisée. En effet, le capital génétique, au sein même d’une espèce, a tendance à diverger de plus en plus si les individus ne se croisent pas dans des groupes. Si des espèces identiques cessent de se retrouver dans les mêmes zones, changent de mode de vie, de mœurs, de manières de se rencontrer, de faire l’amour, etc, alors elles vont cesser de s’interféconder même si l’interfécondation étaient encore génétiquement possible et alors les gènes vont tellement diverger que cela ne sera plus possible par la suite. Aux débuts de la divergence, dans la transition, l’interfécondation est possible. Lorsque des espèces sont en voie de divergence, elles sont interfécondes. La contradiction entre les espèces qui ont une origine commune proche est interne et se développe en marchant. Cette contradiction ne mène pas à la destruction d’une des deux espèces mais à une apparition de nouveauté. Elle est donc dialectique.
Messages
1. L’homme, une espèce révolutionnaire, 11 janvier 2016, 06:48
Repêché au large des îles taïwanaises de Penghu, le fossile d’une mâchoire révèle qu’un groupe inconnu d’hominidés aurait pu côtoyer l’homme moderne il y a encore 10.000 ans. Depuis baptisé Penghu 1 par les paléoanthropologues, cet homo serait âgé de 190 000 ans... à 10 000 ans. Soit une fourchette étonnamment large, qui s’explique par le fait que la mâchoire n’a pas pu être datée au carbone 14, mais simplement via l’analyse d’éléments trace comme le sodium.
2. L’homme, une espèce révolutionnaire, 11 janvier 2016, 06:48, par R.P.
Lire aussi :
Pourrait-il y avoir une autre espèce d’homo qui apparaisse ?
3. L’homme, une espèce révolutionnaire, 17 avril 2016, 07:45, par R.P.
François-Xavier Chauvière dans « Chasseurs-cueilleurs » :
« L’Homme moderne (Cro-Magnon), arrivé en Europe il y a environ 40.000 ans, est souvent considéré comme la seule espèce humaine apte à avoir réalisé des industries sur os, ivoire ou bois de renne. Mais Neandertal, son contemporain pour un temps, peut revendiquer une contribution notable à la production d’objets techniques ou symboliques en matières dures animales. En outre, des découvertes ou réexamens récents d’artefacts mis au jour sur le continent africain viennent relancer le débat sur une probable utilisation technique de la matière osseuse pour des périodes beaucoup plus anciennes. »
4. L’homme, une espèce révolutionnaire, 9 juin 2016, 07:11
L’énigmatique homme de Florès avait probablement des ancêtres, de petits hominidés présents sur la même île il y a 700.000 ans.
De petits hominidés qui vivaient il y a 700.000 ans étaient probablement les ancêtres de l’énigmatique homme de Florès qui vivait sur la même île indonésienne, ont annoncé des archéologues.
Surnommé le "hobbit". Deux études publiées mercredi dans la revue britannique Nature devraient mettre fin à certaines controverses vieilles de plus de dix ans autour des origines de l’homme de Florès, surnommé le "hobbit" en raison de sa petite taille d’environ 1m pour 25 kg. "Ces étranges hominidés étaient donc déjà présents sur l’île il y a 700.000 ans", explique à Yousuke Kaifu, du Musée national de la nature et des sciences d’Ibaraki au Japon. "J’étais stupéfait quand j’ai vu ces nouveaux fossiles" ajoute-t-il.
5. L’homme, une espèce révolutionnaire, 14 juin 2016, 16:32, par Robert Paris
« Nous n’avons pas une très bonne opinion de nos proches parents et nous ne voulons à aucun prix leur ressembler. Les néandertaliens passent pour être arriérés, voûtés, qui s’engouffrent dans leurs cavernes ou en surgissent au pas de charge en traînant leur femme par les cheveux. Le plus récemment découvert, le petit « hobbit » de l’île de Flores en Indonésie, a été déclaré microcéphale et peut-être même « crétin »… Dans les combats internes sans merci dont ils sont coutumiers, les anthropologues évaluent encore les données sur l’homme de Flores, mais la situation des néandertaliens s’éclaircit de jour en jour. Le cliché traditionnel qui faisait d’eux des brutes épaisses n’a jamais été bien convaincant, puisque leur cerveau était plus gros que le nôtre… Lorsque les premiers humains sont sortis d’Afrique, ils ont trouvé des proches parents déjà présents plus au nord depuis deux cent cinquante mille ans, et infiniment mieux adaptés au froid glacial. Au lieu de les vaincre par la conquête, comme dit la version officiellle, peut-être avons-nous établi des liens d’amitié avec ces nordiques. Il est sûr que les hommes ont été séduits par les néandertaliennes, que les femmes ont eu un faible pour les néandertaliens et vice versa, car on estime que jusqu’à 6% de l’ADN des membres non africains de notre espèce viennent des néandertaliens. Ce croisement a probablement beaucoup renforcé notre système immunitaire. Nos frères nordiques enterraient leurs morts, savaient fort bien fabriquer des outils, entretenaient le feu et prenaient soin des infirmes comme les premiers humains. »
Frans de Waal dans « Le bonobo, dieu et nous »
6. L’homme, une espèce révolutionnaire, 15 janvier 2017, 07:17, par R.P.
Les cathos ne supportent pas qu’il y ait eu des tas d’espèces d’hommes !!!
Lire dans "La Croix" !!!!
1. L’homme, une espèce révolutionnaire, 24 février 2017, 07:16, par alias
ne pas confondre prudence scientifique et argumentaire idéologique. Je ne vois pas dans cet article de ce journal une position idéologique mais plutôt un débat entre chercheurs.
2. L’homme, une espèce révolutionnaire, 24 février 2017, 08:41, par Robert Paris
Pour la caractérisation de l’article scientifique, tu as absolument raison. Pour le choix du journal de mettre en avant cette position là et pas l’autre, c’est idéologique, même si c’est défendable.
7. L’homme, une espèce révolutionnaire, 24 février 2017, 08:46, par Robert Paris
Il ne faut pas oublier à quel point la découverte d’un grand nombre d’espèces d’hommes qui ont coexisté pose de problème à la version de la Bible et à la conception d’une seule création divine de l’homme !
8. L’homme, une espèce révolutionnaire, 4 septembre 2019, 07:33
Stephen Jay Gould :
« Voici quelques centaines de milliers d’années il existait une demi-douzaine d’espèces d’êtres humains. Voici seulement 30 000 ou 40 000 ans il en existait peut-être encore trois, avec Neandertal en Europe et Homo erectus en Asie. Aujourd’hui nous nous débrouillons bien, nous sommes nombreux, mais nous sommes tout seuls... »
9. L’homme, une espèce révolutionnaire, 30 janvier 2020, 07:25, par A. Mantoux
L’homme qui apparaissait autrefois comme une seule espèce ou comme une convergence change complètement et apparait comme l’image même du buisson évolutif !!!
La décennie a été marquée par de nombreux progrès concernant la compréhension de nos origines : l’établissement de nouvelles dates sur des fossiles connus, la découverte de crânes fossilisés étonnamment complets ainsi que l’ajout de multiples branches inédites. En 2010, l’explorateur itinérant National Geographic Lee Berger annonce la découverte d’un de nos lointains ancêtres, Australopithecus sediba. Cinq ans après, il révèle que les fossiles d’une nouvelle espèce ont été mis au jour au sein du « berceau de l’humanité », un réseau de grottes karstiques d’Afrique du Sud : Homo. naledi, un hominidé dont l’anatomie en « mosaïque » s’apparente aussi bien à celle de l’homme moderne qu’à celle de ses cousins bien plus anciens. Une étude complémentaire a également révélé le très jeune âge de H. naledi, qui vivait il y a 236 000 à 335 000 ans.
L’Asie a été le lieu de nombreuses découvertes majeures. En 2010, une équipe dévoile que l’ADN identifié à partir d’une phalange d’un ancien habitant de Sibérie ne ressemble à celui d’aucun autre être humain moderne ; il s’agit de la première preuve d’une mystérieuse lignée connue aujourd’hui sous le nom de Dénisoviens. En 2018, des outils de pierre remontant à 2,1 millions d’années sont découverts en Chine, indiquant l’arrivée de fabricants d’outils en Asie des centaines de milliers d’années plus tôt que l’on ne le pensait. En 2019, des chercheurs ont retrouvé des fossiles de Homo luzonensis aux Philippines, une nouvelle espèce humaine semblable à Homo floresiensis, l’Homme de Florès. Enfin, la découverte d’outils de pierre sur l’île des Célèbes antidate l’arrivée de l’Homme moderne, suggérant ainsi la présence d’un troisième hominidé insulaire encore non identifié en Asie du Sud-Est.
La lignée de l’Homme de Denisova, à elle seule, pourrait représenter trois espèces humaines)
10. L’homme, une espèce révolutionnaire, 17 décembre 2020, 05:47, par anonyme
L’homme, une espèce à part ?
https://www.youtube.com/watch?v=stCxLxBMjYA
11. L’homme, une espèce révolutionnaire, 22 juin 2021, 06:06, par A. Mantoux
Une publication, survenue dans la revue américaine Proceedings of the National Academy of Sciences (PNAS) le 15 juin, bouleverse aujourd’hui l’état des connaissances dans le domaine préhistorique. Elle documente en effet la cohabitation, en un même lieu, de nos lointains ancêtres Sapiens avec des hommes (et des femmes) de Neandertal. De cette rencontre entre ces deux lignages de l’humanité résulterait la transformation des techniques de taille du silex, intervenue il y a 50 000 ans, dans cette région.
Selon les conclusions de l’article, cosigné par une dizaine de scientifiques israéliens et français (parmi lesquels Elisabetta Boaretto, Marion Hernandez, Mae Goder-Goldberger, Vera Aldeias, Lior Regev, Valentina Caracuta, Shannon P. McPherron, Jean-Jacques Hublin, Steve Weiner et Omry Barzilai), cette révolution technologique se serait produite de manière plus rapide que ce que l’on pensait jusque-là. Elle aurait permis l’adoption de nouvelles techniques de chasse. Les pointes taillées permettant la mise au point d’armes de jet : javelots, puis flèches.
Le site exploré est localisé à Boker Tachtit, non loin de la localité de Sde Boker, au centre du Néguev. Il a été découvert dans le courant des années 1980. Une campagne de fouille organisée entre 2013 et 2015 à l’initiative du centre Weizmann a permis d’exhumer sur place un important mobilier de pierres taillées, mais aussi des fragments de charbon de bois dont la datation au carbone 14 atteste une occupation humaine il y a 50 000 ans. Si aucun reste humain n’a été retrouvé localement, c’est la diversité des techniques de taille qui démontre la coexistence sur place de ces souches humaines cousines.
Les Neandertaliens seraient arrivés dans la région, en provenance du nord, il y a 70 000 ans. L’arrivée de Sapiens, depuis le continent africain, serait de son côté intervenue il y a 200 millénaires. Ce qui permet de penser que les confrontations entre les deux lignées sont antérieures à celles attestées à Boker Tachtit. Les uns et les autres coexistant et probablement interagissant.
Mais c’est une vague migratoire plus tardive, remontant par la péninsule arabique ou le long du Nil, autour de 50 000 ans avant notre ère, qui aurait permis à la fois le métissage génétique, que postule la théorie de « l’origine africaine moderne », ainsi que l’introduction d’une nouvelle culture, indique Elisabetta Boaretto.
https://www.lepoint.fr/culture/l-incroyable-histoire-de-la-femme-percutee-par-une-meteorite-23-03-2021-2418943_3.php
12. L’homme, une espèce révolutionnaire, 27 juin 2021, 05:40, par A.M.
Les généalogies de l’homo sapiens et néanderthal sont bouleversées
https://www.futura-sciences.com/planete/actualites/ancetres-humain-neandertal-detrone-homo-longi-alias-dragon-man-serait-notre-plus-proche-cousin-92141/
https://hitek.fr/actualite/deux-decouvertes-majeures-embranlent-ce-qu-on-pensait-savoir-de-neandertal_29612
13. L’homme, une espèce révolutionnaire, 2 juillet 2022, 08:18, par Luc Garde
Encore une découverte révolutionnaire sur l’évolution de l’homme
https://www.lemonde.fr/sciences/article/2022/06/27/en-afrique-du-sud-des-australopitheques-toujours-plus-vieux_6132253_1650684.html