
Accueil > 02 - Livre Deux : SCIENCES > Evolution ou révolution des espèces ? > Qu’est-ce que l’évolution des espèces « en buisson » et qu’est-ce que cela (...)
Qu’est-ce que l’évolution des espèces « en buisson » et qu’est-ce que cela change de fondamental ?
mardi 1er octobre 2019, par


« L’évolution de la vie à la surface de la planète est conforme au modèle du buisson touffu doté d’innombrables branches et continuellement élagué par le sinistre sécateur de l’extinction. Elle ne peut du tout être représentée par l’échelle d’un inévitable progrès. »
Stephen Jay Gould
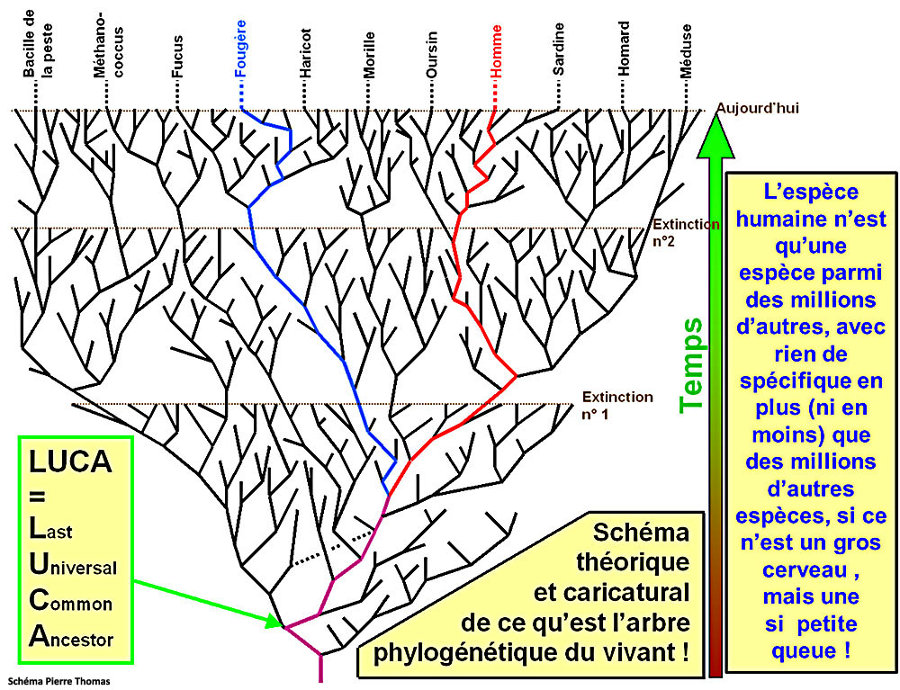


La vie représentée en buisson, dessinée par Darwin dès 1837 dans son « carnet B »

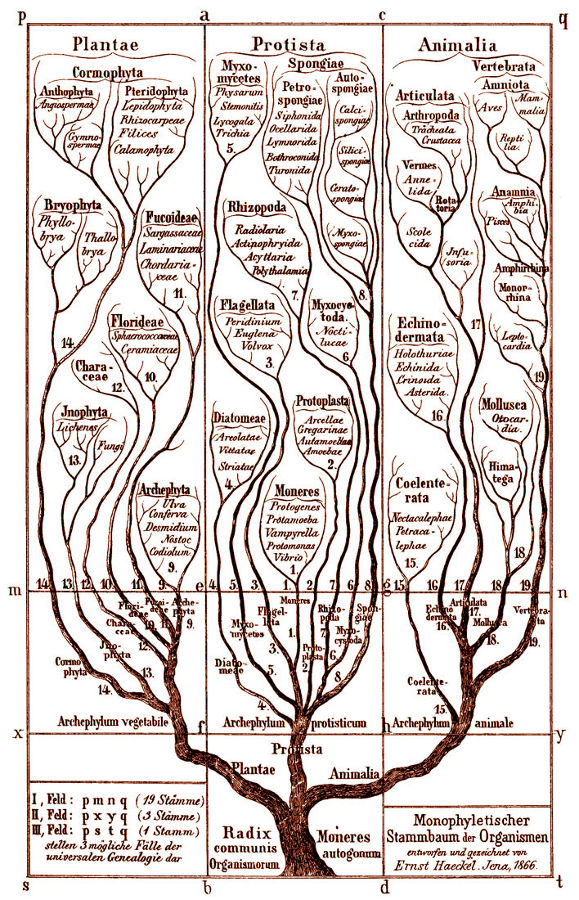
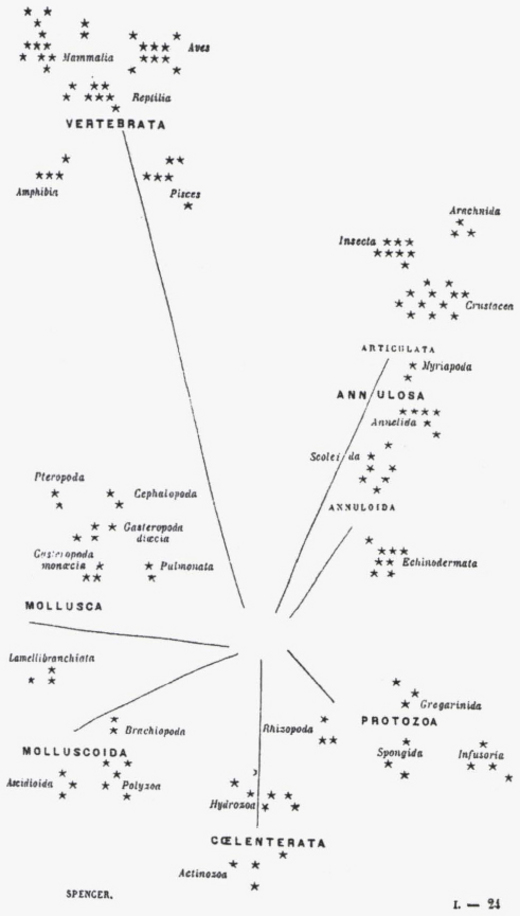
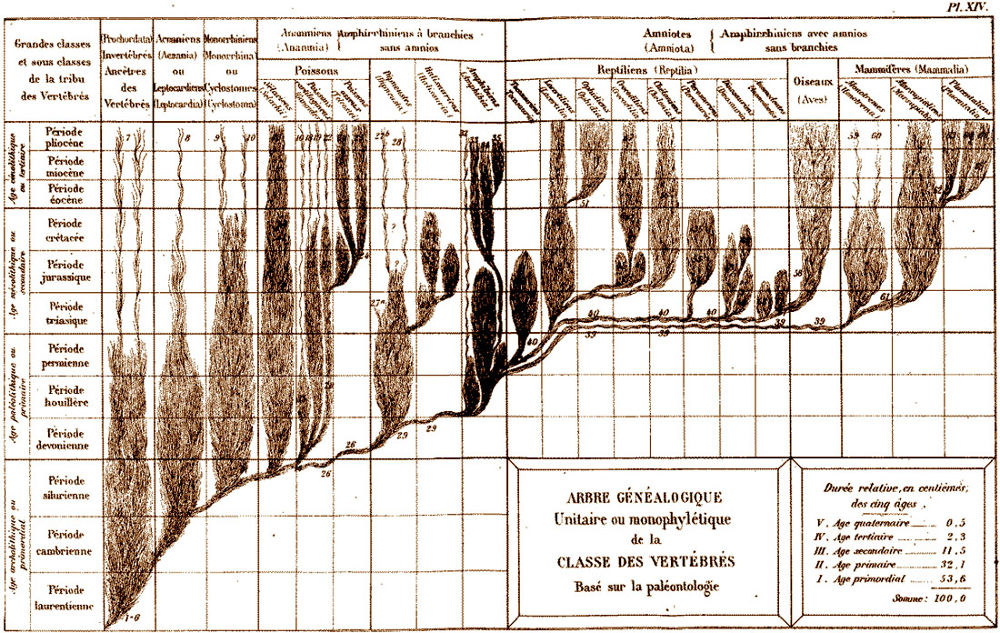
Les espèces vivantes, un buisson tantôt foisonnant de diversité et tantôt massivement élagué

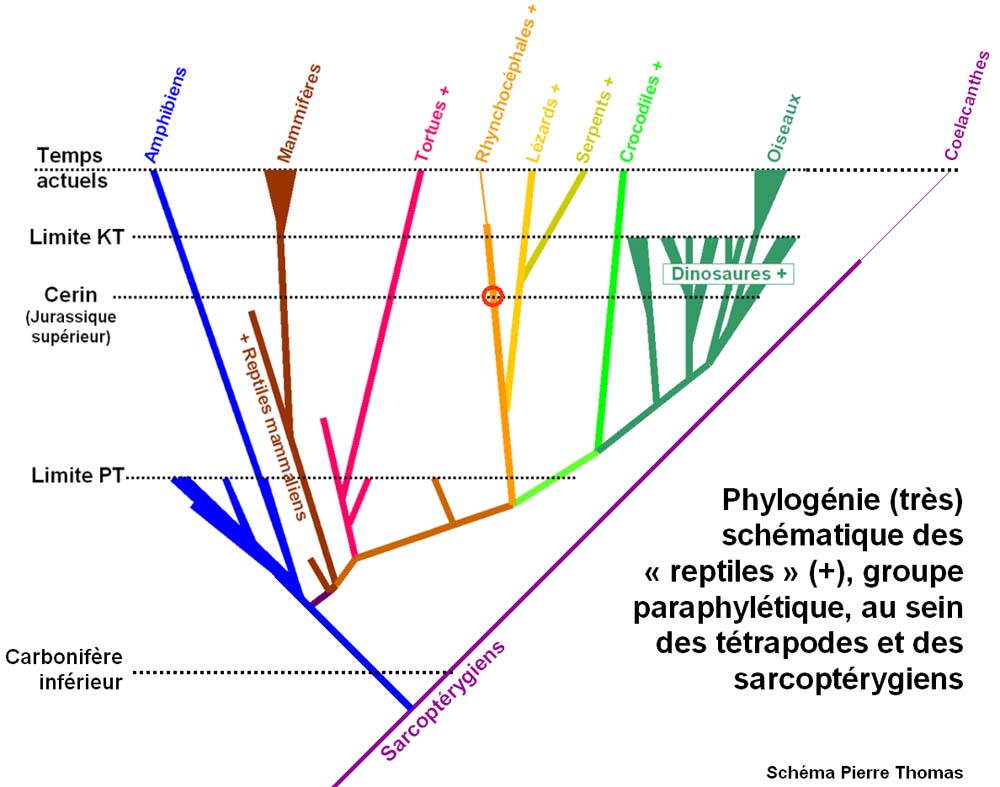
Dans le schéma du buisson, contrairement à l’ancien "arbre de l’évolution", toutes les espèces d’une même époque sont aussi éloignées des origines et donc aussi "évoluées"


Qu’est-ce que l’évolution des espèces « en buisson » et qu’est-ce que cela change de fondamental ?
On nous avait longtemps parlé de l’ « arbre de l’évolution » et cela semblait indiquer une évolution linéaire de bas en haut et on choisit de nos jours de souligner plutôt une image du « buisson ». Mais qu’est-ce que cela change ? Eh bien, dans un arbre, toutes les branches grandissent, les grosses et les petites. Pour le buisson, l’image sous-entend une série de branches toutes égales qui sont taillées au hasard d’un côté ou d’un autre, les branches coupées (par des extinctions locales ou générales) s’interrompant définitivement sans qu’aucune des petites branches soit plus centrale, plus durable ou plus prédictivement porteuse d’avenir. L’image du buisson récuse la notion de progrès et même de progression. Les brindilles survivantes reforment un nouveau buisson par la diversification et, à nouveau, les extinctions taillent le buisson. Cette image signifie que l’on voit alterner des périodes de diversification et des périodes d’extinction. Ce n’est pas un arbre qui est sans cesse en train de croitre et de grandir vers le haut. Ce sont des successions d’explosions de diversité et de taillage de haies de destructions massives d’un grand nombre d’espèces qui dessinent l’arborescence de la vie dans une forme très particulière et non l’évolution conçue comme un progrès général vers le haut dont les meilleurs éléments seraient les seuls conservés. On constate en effet qu’un très grand nombre d’espèces sont nées en même temps et sont disparues dans des périodes de disparitions de masse. Cela donne dont une idée très différente de ce que l’on a appelé faute de mieux « l’évolution » mais qui ne doit surtout pas être conçu comme « évolution vers » les espèces qu’il y a aujourd’hui. Le buisson renvoie à la notion des « équilibres ponctués » de Stephen Jay Gould et écarte les conceptions adaptationnistes du progrès, téléologiste et gradualiste de la transformation au long de la descendance. Le buissonnement est touffu mais il est coupé, à nouveau repart en touffe et, à nouveau, est durement coupé. C’est une conception tout à fait nouvelle. Les rescapés de la vie sont des espèces qui ont des ancêtres communs plus ou moins lointains mais qui n’appartiennent pas à une même branche. La vision linéaire, continue et progressive a été abandonnée. La directivité de l’arbre ne semble plus la meilleure image du réel. L’image du buisson ne semble favoriser aucune branche, aucun embranchement. Elle représente toutes les espèces existant actuellement comme ni plus ni moins favorisées ou évoluées les unes que les autres, le buisson étant circulaire et indiquant seulement l’écoulement du temps. C’est cela qui nous amène à représenter plutôt le changement comme révolutions que comme évolution...
« L’histoire de la vie ressemble à un gigantesque élagage ne laissant survivre qu’un petit nombre de lignées, lesquelles peuvent ensuite subir une différenciation ; mais elle ne ressemble pas à cette montée régulière de l’existence, de la complexité et de la diversité, comme on le raconte traditionnellement… Pour les spécialistes, l’évolution est une adaptation aux conditions changeantes de l’environnement et non pas un progrès… L’évolution de la vie à la surface de la planète est conforme au modèle du buisson touffu doté d’innombrables branches et continuellement élagué par le sinistre sécateur de l’extinction. Elle ne peut du tout être représentée par l’échelle d’un inévitable progrès. »
« La vie est belle », Stephen Jay Gould
« L’évolution procède par embranchements, et non par métamorphose d’une forme à une autre, l’ancien disparaissant dans le triomphe du nouveau… En outre, la plupart des nouveautés, du moins à leurs débuts, croissent comme de minuscules brindilles sur des buissons vigoureux et persistants, et non comme des réalisations plus sophistiquées d’ancêtres qui ont donné leur maximum à un organisme qui transcende la médiocrité de leurs êtres. Les amphibiens et leurs descendants ont certes bien réussi sur la terre ferme, mais les nageoires sont supérieures aux pieds dans le buisson des vertébrés ; la majorité de ses pousses (espèces) y sont des poissons. Je ne nie pas le succès actuel des humains et la nouveauté intéressante qu’ils représentent. »
Stephen Jay Gould dans « Cette vision de la vie »
« L’évolution se fait généralement par « spéciation » (isolement d’un lignage qui perd tout contact avec le stock parental) et non par transformation lente et continue de ce stock parental. Des épisodes répétés de spéciation produisent un buisson. Les « séquences » d’évolution ne sont pas les barreaux d’une échelle, mais un chemin sinueux, labyrinthique, conduisant de branche en branche, de la base du buisson à l’espèce qui subsiste actuellement au sommet. »
Stephen Jay Gould dans « Darwin et les grandes énigmes de la vie »
L’hominisation passe ainsi d’une conception simple et linéaire à une conception beaucoup plus complexe, buissonnante en référence au buisson, dessiné par Darwin dès 1837 dans le « carnet B », puis explicitement proposé par Gould en 1979 (p.63) : « Nous ne sommes pas le sommet d’une échelle destinée, dès le départ, à produire l’Homo sapiens. Nous sommes tout simplement la seule branche restante d’un buisson jadis luxuriant ». Ainsi, après la confrontation à la réalité de formes d’hominidés contemporaines, la ligne qui menait à l’homme s’est transformée en rameau, puis le rameau en buisson et actuellement c’est un buisson où l’apparition des caractères qu’on pensait propres à la lignée humaine peut se faire dans des rameaux non en relation directe avec l’homme. La bipédie aurait pu apparaître à plusieurs reprises, ainsi que le développement du cerveau. Il est difficile dans ce contexte de proposer un arbre phylogénétique pertinent et certains en viennent à proposer d’abandonner cette ambition : « la prudence voudrait sans doute qu’on cesse de construite des arbres d’évolution et qu’on attende que tout ce savoir soit enrichi, précisé et élagué. Un arbre ne fait rien d’autre que cacher la forêt... » (Cohen, 2001, p.33-37)… L’évolution menant du singe à l’homme n’a pas été cette échelle orientée que certains se plaisaient à imaginer ; elle donne plutôt l’image d’un buisson, d’un arbre plein de rameaux divergents, de tentatives ratées et de bifurcations. »
Quessada Marie Pierre
« Le développement de l’art paléolithique s’est probablement fait de façon semblable à l’évolution elle-même : non pas sur le modèle d’une ligne droite ou d’une échelle, mais de façon bien plus compliquée, évoquant la croissance complexe d’un buisson avec des branches parallèles et des quantités de ramifications ; non pas sur le mode du changement lent et graduel, mais sur celui de « l’équilibre ponctué », avec des phases brèves et occasionnelles de réussite éclatante. »
Bahn et Vertut (1998)
« En outre, la plupart des nouveautés, du moins à leurs débuts, croissent comme de minuscules brindilles sur des buissons vigoureux et persistants, et non comme des réalisations plus sophistiquées d’ancêtres qui ont donné leur maximum à un organisme qui transcende la médiocrité de leurs êtres… L’arbre de vie qui imaginait une progression graduelle des phyla à partir de formes simples (comme les éponges) vers une vie plus complexe (comme les vers), puis vers les créatures à coquilles (comme les mollusques), a été remplacé par le " buisson " de vie dans lequel les éponges, les vers, les mollusques et tous les autres 34 phyla sont apparus simultanément. Chacune de ces lignes issues du buisson ont ensuite développé une myriade de variations, mais les variations sont toujours restées à l’intérieur du plan fondamental des corps. »
Gerald Schroeder, dans "L’évolution, rationalité ou hasard ?"
« Nous avons tendu jusqu’à présent à voir l’histoire de notre lignée comme moins touffue qu’elle n’a réellement été. En même temps, nous avons toujours tendu à voir notre propre espèce comme plus centrale dans l’évolution de notre famille qu’elle ne peut l’être, étant donné qu’elle ne représente en fait qu’une brindille terminale parmi d’autres au sein d’un gros buisson (mais il est vrai la seule survivante aujourd’hui). »
Ian Taterstall
« Une approche étymologique (A.Rey 1998) indique que, dès les premiers emplois connus du terme buisson, ce mot « désigne un bouquet d’arbrisseaux sauvages et rameux ». A contrario, la lignée « désigne seulement la descendance d’une personne », ce qui est classiquement représenté par un « arbre » généalogique. Une approche épistémologique suggère que les enjeux du remplacement du terme lignée par celui de buisson ne sont pas que langagiers. Tout d’abord, le sens commun de lignée, et d’arbre généalogique, s’oppose au sens scientifique de « lignée évolutive » et « d’arbre phylogénétique », ce qui risque d’introduire deux obstacles épistémologiques, qui sous-tendent les commentaires qui suivent. D’une part une confusion entre le temps historique récent d’une généalogie et le temps géologique de l’évolution ; d’autre part une confusion entre les liens directs de parenté entre individus dans une généalogie, et les liens indirects et hypothétiques entre espèces différentes dans une phylogenèse. La succession des espèces durant les temps géologiques se schématise par une représentation où figure la répartition des espèces fossiles en fonction de l’échelle stratigraphique du temps géologique. La présence de chaque espèce est indiquée par un trait vertical d’autant plus long que cette espèce a été retrouvée sous forme de fossiles durant une période géologique plus importante. Il s’agit de faits. La métaphore la plus courante indique que chaque espèce fossile correspondrait à une feuille d’arbre dont les branches, ramifications, troncs, n’ont pas laissé de traces. Ce type de schéma ne représente que ces « feuilles », sans lien entre elles. Etablir ces liens, sous forme toujours hypothétique, est l’objectif de la phylogénie. Ces liens résultent d’une part de la comparaison entre les espèces (au niveau des organes, des molécules, etc), d’autre part d’hypothèses sur les mécanismes évolutifs, telles que le principe de parcimonie pour les cladogrammes (Tassy 1986, Mayr 1996). Les comparaisons entre espèces (actuelles ou fossiles) permettent ainsi d’établir « des arbres phylogénétiques » qui les relient en fonction de leurs plus ou moins grandes ressemblances. « Un noeud (point de rencontre entre une branche mère et deux branches fille) n’exprime pas un spéciation mais une parenté fondée sur le partage de caractères uniques. En d’autres termes, le cladogramme dit « qui partage quoi avec qui » et donc « qui est plus proche parent de qui » et non pas « qui descend de qui » » (Lecointre, 1995). Ces deux modes de schématisation sont donc bien distincts, l’un représentant la présence d’espèces et la durée supposée de leur existence, l’autre des liens possibles, hypothétiques, entre ces espèces, sans datation de ces liens. Ils peuvent être superposés sur un schéma mixte qui tente de représenter des liens entre les espèces fossiles disposées en fonction de l’échelle des temps géologiques. Une première différence entre « arbre » et « buisson » peut ainsi être proposée. •En se situant à une origine de la vie supposée unique, les parentés entre toutes les espèces vivantes (actuelles comme fossiles) sont souvent conçues sous la forme d’un « arbre phylogénétique ». Cette construction intellectuelle suppose que cet arbre traverse les temps géologiques, donne des branches qui meurent (les espèces et groupes aujourd’hui disparus) et d’autres qui sont actuellement présentes sur terre. •En se situant dans une tranche de temps précise (récente et actuelle, ou lors d’une période géologique précisément datée), nous sommes confrontés à une coupe horizontale de l’arbre précédent, qui a alors l’aspect buissonnant de plusieurs arbrisseaux juxtaposés (et concurrents) dans un espace précis. Arbres, branches, ramifications, buissons, sont donc autant de métaphores possibles pour désigner des liens hypothétiques entre espèces, alors que le terme « lignée » évoque plus une succession directe entre espèces. Ainsi, tous ces termes cachent des implicites qu’il convient d’identifier.
Gradualisme ou équilibres ponctués ?
S.J. Gould (1979) a proposé l’idée d’une évolution buissonnante, en développant clairement « le conflit entre « échelles » et « buissons » ». La métaphore de l’échelle, couramment employée pour l’évolution de l’homme, « renvoie à la conception populaire selon laquelle l’évolution serait une suite ininterrompue d’ancêtres et de descendants ». Ainsi se succèderaient de façon linéaire différentes espèces d’Homininés, certains étant déjà connus et d’autres correspondant à des « chaînons manquants ». Cette métaphore de l’échelle est aujourd’hui tombée en désuétude, mais a été relayée par celle de « lignée ».Derrière ces métaphores, ce sont deux visions théoriques divergentes qui se manifestent : L’échelle ou la lignée correspondent au modèle de l’évolution dite anagénétique, ou gradualisme, avec modification lente et progressive d’une espèce qui en donne une nouvelle. De plus, cette représentation est souvent finalisée (par exemple la lignée qui aboutit à Homo sapiens). Le buisson correspond au modèle des équilibres ponctués (Elredge & Gould 1972, Devillers 1996). Selon ce modèle, il est improbable de trouver des fossiles correspondant au moment de la formation de l’espèce qui est relativement rapide au sein d’une petite population. Les fossiles ne correspondent donc qu’à des espèces stabilisées et apparemment discontinues. Le premier modèle s’accorde avec le regroupement de nombreux fossiles au sein de la même espèce ou du même genre. Le second modèle, qui correspond à la tendance actuelle, consiste à distinguer un grand nombre d’espèces et de genres qui coexistent. Le nombre d’espèces fossiles décrites a fortement augmenté de 1964 à 2003, ce qui a aussi contribué à la nouvelle schématisation d’une évolution buissonnante de l’espèce humaine (Langlois 2004). L’introduction de la notion d’évolution buissonnante est donc associée à une remise en cause du modèle évolutif gradualiste. Ce premier modèle est lié une conception simplificatrice, finaliste et anthropocentrée de l’évolution alors que le modèle en buisson est associé à une conception contingente de l’évolution humaine.
Lignées (arbres, buissons) ou réseaux ?
La notion d’évolution en mosaïque (introduite par De Beer 1954 : voir Delsol 1996) attire l’attention sur le fait que les organes et les structures d’une lignée n’évoluent pas à la même vitesse : ils évoluent indépendamment les uns des autres.
La notion d’évolution en réseau va plus loin (Bonhomme 1996). Elle s’oppose à la phylogénie dichotomique en branches d’arbre, et propose la possibilité de processus évolutifs « par échanges génétiques secondaires entre taxons ». Ce qui peut venir soit de croisements entre populations géographiquement éloignées, soit de modifications des génomes par d’autres moyens que la reproduction sexuée (virus, transposons). Pour exemple, la théorie polycentriste sur l’apparition des hommes modernes admet des échanges génétiques répétés entre les populations d’Homo erectus de tout l’ancien monde, assurant une continuité génétique et une tendance évolutive commune vers Homo sapiens (Leakey, 1997). »
Quessada Marie Pierre
« C’est un peu la fable de l’arbre et du buisson. L’arbre, c’est la manière dont trop souvent encore, en France ou ailleurs, on présente les origines de l’homme : comme un tronc planté droit, sans presque aucune branche morte, qui aurait vu se succéder les australopithèques, puis Homo habilis, Homo erectus, et enfin Homo sapiens. Le buisson, c’est désormais l’image de l’évolution qui se dégage des découvertes advenues au cours de ces vingt dernières années. Des branches mortes multiples et sans descendance, une évolution qui tâtonne, qui bricole sous l’empire du seul hasard, au point qu’il n’y a pas « naissance » de l’homme, mais « émergence », fruit imprévu d’adaptations contingentes. »
Ian Tatersall
« Appartenir à la lignée humaine dont le seul représentant actuel est l’Homme c’est essentiellement être bipède, avoir la face réduite, une capacité crânienne importante, fabriquer des outils, avoir une pensée consciente associée à un langage articulé. La lignée humaine présente un caractère buissonnant, c’est-à-dire que l’évolution des caractères ne s’est pas faite d’une, manière conjointe et régulière. En effet plusieurs genres et espèces d’hommes présentant des associations variées de caractères plus ou moins évolués, ont coexisté à plusieurs moments. On peut donc admettre que l’évolution progresse selon un mode buissonnant par rameaux divergents. Cependant l’étude de la lignée humaine présente quelques difficultés. En effet les fossiles découverts sont souvent fragmentaires et il est difficile de déterminer à quelle espèce ils appartiennent. Par ailleurs, les découvertes récentes peuvent remettre en cause une phylogénie établie et il n’est pas exclu que la présence uniquement africaine des plus anciens fossiles d’homininés soit due à des conditions de fossilisation exceptionnelles dans la vallée su rift africain. »
Thierry Maulnier
« La lignée humaine – on dit aussi le « genre Homo » – fait figure d’une brindille dans le buissonnement du vivant depuis 3,8 milliards d’années. Parmi les mammifères, et dans le bouquet des primates, cette lignée très courte appartient au petit buisson des singes sans queue – on dit aussi les « Hominoïdes ». »
Boris Valentin
« L’homme fait partie du buissonnement du vivant. En termes biologiques, il n’est pas plus évolué que les autres » espèces. »
Evelyne Heyer
« Le mot même d’évolution est ambigu. Darwin, dans ses premiers écrits, refusa ce mot parce que sa théorie ne contenait aucune notion d’amélioration générale. Il préférait parler de « descendance avec modification ». Finalement, c’est sous l’influence du philosophe Herbert Spencer que ce mot s’est imposé. Il est resté et il est trop tard pour le changer. Ce que nous voyons au cours du temps, c’est l’apparition et la disparition d’espèces qui durent en moyenne 4 millions d’années. Le maximum, 30 millions d’années, ayant été atteint par une certaine crevette. Chaque lignée passe par des phases de buissonnement (multiplication rapide des espèces), de stagnation et de réduction de la population. L’homme est-il alors aussi une espèce menacée d’extinction ? Je ne peux pas faire de prédiction. La seule chose que l’on puisse dire, c’est que toutes les espèces finissent par disparaître. Il y a 2 à 3 millions d’années, coexistaient une demi-douzaine d’espèces d’australopithèques, dont une a donné naissance au genre Homo. A l’époque florissant, puisqu’il y a eu habilis, ergaster, rudolfensis, erectus" Aujourd’hui, il n’y a plus qu’une seule espèce. Nous ne sommes pas les seuls dans ce cas et, en général, on les appelle des « échecs » de l’évolution : par exemple le tuatara, reptile de Nouvelle-Zélande, ou encore le pangolin (un mammifère édenté, ndlr). Et même les chevaux, que l’on présente souvent comme un triomphe de l’évolution, ne sont plus représentés que par huit espèces. »
Stephen Jay Gould, interview de septembre 1997
« Pour ne pas se perdre parmi tous les prétendants au titre de l’ancêtre de l’homme, il est important de comprendre en amont le principe de buissonnement. Contrairement à ce que l’on a longtemps cru, l’évolution n’est pas linéaire et peut donner naissance à de nombreuses espèces issues d’un même ancêtre : le Dernier Ancêtre Commun (le DAC). Quel est par exemple le DAC du cheval et de l’âne ? Celui du perroquet et de l’aigle ? Il s’agit de la dernière étape avant de remonter le temps et de se retrouver aux débuts de la vie sur Terre. »
Jacques Pernaud
Les thèses fondamentales de l’équilibre ponctué de Stephen Jay Gould :
« La théorie de l’Equilibre ponctué ne concerne pas toutes les formes de changement rapide en biologie, se produisant à n’importe quelle échelle ou à n’importe quel niveau. Elle porte sur l’apparition et le déploiement des espèces à l’échelle des temps géologiques. D’autres phénomènes, prenant place à d’autres échelles, sont aussi de type ponctuationniste : c’est le cas, par exemple, des extinctions de masse catastrophiques planétaires déclenchées par la collision de la Terre avec des météorites…
La théorie de l’équilibre ponctué essaie d’expliquer le rôle macroévolutif des espèces et de la spéciation dans le cadre des temps géologiques. Les phases de changement rapide et de stabilité qu’elle décrit se rapportent à l’histoire des espèces individuelles ; et les rythmes et les types de changement qu’elle prend en compte concernent le déploiement de ces histoires individuelles dans un domaine qui nous est familier, celui du « temps profond », autrement dit des temps géologiques, à l’échelle desquels la durée de vie humaine est un infiniment petit absolument impossible à mesurer, et la durée de l’histoire entière de la civilisation humaine par rapport à celle de la phylogenèse des primates comparable à la durée d’un battement de paupière par rapport à celle de la vie humaine…
La conception fondamentale de l’équilibre ponctué comporte trois notions dont il est nécessaire de définir le sens de façon opérationnelle et précise : la stase, la ponctuation et la fréquence relative dominante…
La stase ne signifie pas une « stabilité de granite », autrement dit une totale invariance des valeurs moyennes de tous les traits tout le temps. Dans le contexte macroévolutif de l’équilibre ponctué, il est nécessaire de savoir, par-dessus tout, si le changement morphologique tend ou non à progresser de façon cumulative au long de l’existence géologique d’une espèce et, si oui, quelle fraction de la différence moyenne entre une espèce ancestrale et une espèce descendante peut être attribuée au changement cumulatif subi par l’ancêtre au cours de son évolution anagénétique…
Puisque l’existence de la stase se fonde sur des données, tandis que la ponctuation correspond généralement à une transition qu’il est impossible de détailler par le moyen classique de la distribution des fossiles au fil du temps géologique, il est nécessaire de formuler une définition appropriée de la rapidité… En première approximation, la durée correspondant à un plan de stratification définit la limite pratique imposée à la possibilité de distinguer l’un de l’autre deux phénomènes survenus dans le temps géologique. Tout épisode de spéciation qui se produit en un intervalle de temps correspondant à la durée généralement nécessaire à la réalisation d’un plan de stratification se trouvera ramassé en une seule couche stratigraphique mince, représentant un « instant » à l’échelle des temps géologiques, et ne pourra donc généralement pas être analysé de façon détaillée…
Il faut donc définir les ponctuations par rapport à la durée de la stase des espèces qui en sont issues : car la théorie de l’équilibre ponctué, envisageant le déroulement dans le temps de phases de durée différente, soutient que les espèces acquièrent réellement leurs caractères distinctifs au « moment de leur naissance », et qu’elles le gardent ensuite en stase durant la durée de leur longue existence géologique. Ces questions de durées relatives jouent un rôle important dans la définition des espèces en tant qu’individus darwiniens…
La théorie de l’équilibre ponctué soutient, et c’est sa thèse la plus importante, que cette forme d’évolution est dominante en termes de fréquence relative, et ne dit donc pas seulement que ce phénomène existe… La théorie de l’équilibre ponctué n’affirme pas simplement que ce phénomène existe, mais avance la thèse plus ambitieuse selon laquelle il joue un rôle dominant en tant que forme prise par la macroévolution dans le cadre des temps géologiques…
Eldredge et moi-même avons forgé le terme d’équilibre ponctué dans une communication orale originellement présentée lors d’un colloque intitulé « Modèles en paléobiologie », qui s’est déroulé en 1971 dans le cadre de la réunion annuelle de la Société géologique d’Amérique… C’est ce qui a donné notre article original sur l’équilibre ponctué – Eldredge et Gould, 1972… Un problème nous avait particulièrement agacés, c’était le difficulté de mettre la main sur des séquences gradualistes dans les archives fossiles, afin de pouvoir leur appliquer des techniques statistiques et autres méthodes quantitatives…
Avant que nous ayons proposé la théorie de l’équilibre ponctué, la plupart des paléontologistes pensaient que, pour sa plus grande part, le changement évolutif procédait sur le mode de l’anagenèse, autrement dit par transformation continue au cours du temps d’une population donnée dans son ensemble. C’est pourquoi la plus grande partie des discussions en paléontologie concernant les espèces tournait alors autour d’une question litigieuse, nommément ce qu’on appelle le problème de l’espèce en paléontologie, lequel a sans cesse été remis sur le chantier dans notre littérature (voir Imbrie 1957 ; Weller, 1961 ; McAlester, 1962 ; Shaw, 1969) et a même conduit à des colloques entiers consacrés aux solutions éventuelles (voir Sylvester-Bradley, 1956).
Ce prétendu problème relève plus de la théorie abstraite et des définitions que des faits observés, car il s’appuie sur l’idée selon laquelle l’anagenèse serait dominante dans la réalité. En tout cas, c’est bien dans le cadre du gradualisme qu’il se pose, parce qu’un vrai continuum ne peut être divisé avec certitude absolue en segments auxquels peuvent être attribués des noms distincts. Si une population A change si complètement par le biais de l’anagenèse que l’on se sent obligé de donner à la population en résultant une nouvelle dénomination linnéenne (espèce B), où faut-il placer le point de démarcation entre A et B ? Toute limite de ce genre ne peut être qu’arbitraire…
La théorie de l’équilibre ponctué a adopté une approche radicalement différente : elle a admis que, dans les conditions ainsi définies, il était, en effet, impossible de trouver une limite nette de séparation entre espèce parentale et espèce descendante ; mais elle a alors nié la prémisse empirique précise selon laquelle les nouvelles espèces naissent généralement (ou même souvent) par le biais de l’anagenèse gradualiste.
Au contraire, Eldredge et moi-même avons soutenu que la vaste majorité des espèces naissent à l’issue d’une scission, et que le rythme normal de la spéciation, tel qu’il s’exprime dans les temps géologiques, conduit à l’apparition d’une nouvelle espèce en un instant géologique, celles-ci persistant ensuite de façon prolongée en stase…
Bien entendu, les partisans du gradualisme ne niaient pas que la spéciation se produisit souvent pas scission. Mais ils ne pensaient pas que ce processus de scission jouât un rôle quelconque dans la macroévolution, cela pour trois raisons. Premièrement, ils concevaient la spéciation seulement comme un mécanisme engendrant de la diversité, non comme un facteur de changement de la morphologie moyenne au sein d’un clade (autrement dit, comme un facteur responsable de ce phénomène macroévolutif crucial que sont les tendances évolutives)…
Deuxièmement, ils n’accordaient que peu de place au processus de spéciation (naissance des espèces résultant d’une scission) par rapport à l’anagenèse dans l’ensemble du changement évolutif…
Troisièmement, lorsqu’il leur arrivait d’évoquer un phénomène de spéciation par scission, ils dépeignaient ce processus comme deux épisodes d’anagenèse au-delà de la scission, se déroulant chacun selon le rythme lent caractéristique de cette forme d’évolution. Ainsi, ils n’apercevaient rien de fondamentalement différent dans le changement évolutif réalisé par spéciation. En raison de certains accidents de l’histoire, soutenaient-ils, une population se scindait en deux unités séparées, chacune évoluant alors selon le mode anagénétique habituel.
La théorie de l’équilibre ponctué, d’un autre côté, propose que le rythme de réalisation de la spéciation, apprécié à l’échelle des temps géologiques, diffère radicalement de celui de l’anagenèse gradualiste…
Premièrement, la théorie de l’équilibre ponctué assure l’expansion hiérarchique de la théorie sélectionniste au niveau de l’espèce, ce qui permet de dépasser le choix qu’avait fait Darwin de restreindre dans les faits les mécanismes causals de l’évolution au seul niveau des organismes.
Deuxièmement, en définissant les espèces comme les unités fondamentales (ou les atomes) de la macroévolution (autrement dit, comme les entités stables – des individus darwiniens – et non comme des parties arbitrairement délimitées au sein de processus continus), la théorie de l’équilibre ponctué interdit d’expliquer la totalité des aspects à grande échelle de l’évolution par simple extrapolation des résultats de la microévolution obtenus sur des populations locales, à l’échelle du temps humain et au niveau organismisque ou même à des niveaux inférieurs…
En ce qui concerne le rythme évolutif, les équilibres ponctués renversent la vision fondamentale. Il faut abandonner la notion d’un changement constant, qui opérerait sur un rythme bien net et important, et caractériserait l’état normal d’une entité en train d’évoluer. Il faut donc procéder à une révision et voir désormais le changement évolutif sous la forme d’une série de rares épisodes, de durée brève par comparaison à celle des périodes de stase qui les séparent. La stabilité est désormais l’état normal d’un lignage, tandis que le changement est à présent conçu comme un phénomène se produisant rarement et dans une période de temps limitée, mais rendant compte néanmoins de la phylogenèse par le biais d’une série d’épisodes additionnés au cours du temps.
Cette révision fondamentale dans la façon d’appréhender les choses peut trouver des échos dans toutes sortes de domaines, des plus immédiatement pratiques jusqu’aux plus globalement philosophiques.
Dans un cadre concernant plus immédiatement la biologie, la même révision fondamentale dans la façon d’appréhender les choses conduit inéluctablement à mettre l’accent davantage sur le hasard et la contingence que sur la prédictibilité fondée sur l’extrapolation : car l’état ordinaire de stase ne permet guère de savoir quand et comment va se produire la ponctuation suivante, tandis que la nature fractale du gradualisme conduit à penser que les causes du changement à n’importe quel moment peuvent, par extrapolation, permettre de prédire et d’expliquer les vastes effets observés par l’accumulation des petits changements au fil des longues durées…
De nombreuses recherches menées dans le domaine des sciences sociales, des arts et de la littérature, la théorie des révolutions scientifiques de Thomas Kuhn étant la plus connue et la plus influente, ainsi que de nombreux événements de la fin du XXe siècle se sont combinés pour susciter une prise de conscience aigüe du caractère insatisfaisant du gradualisme et de l’acceptabilité générale du changement ponctuationniste, au point d’en faire presque une orthodoxie…
Les données de l’observation scientifique ont aussi contribué au développement de ce mouvement général en fournissant des modèles et des vérifications pratiques à différents niveaux d’analyse et pour plusieurs systèmes. Dans ce cadre, la paléontologie a fourni un apport bien connu, celui d’une théorie d’extinction de masse catastrophique…. »
« La structure de la théorie de l’évolution » de Stephen Jay Gould
« Toutes les grandes théories de la spéciation s’accordent à reconnaître que la divergence s’effectue rapidement au sein de populations très réduites. (...) Le processus (de spéciation) peut prendre des centaines voir des milliers d’années. (...) Mais mille ans, ce n’est qu’un infime pourcentage de la durée moyenne d’existence des espèces invertébrées. (...) Eldredge et moi faisons référence à ce mécanisme sous le nom de système des équilibres ponctués. (...) Si le gradualisme est plus un produit de la pensée occidentale qu’un phénomène de nature, il nous faut alors étudier d’autres philosophies du changement pour élargir le champ de nos préjugés. Les fameuses lois de la dialectique reformulées par Engels à partir de la philosophie de Hegel, font explicitement référence à cette notion de ponctuation. Elles parlent par exemple de ‘’ la transformation de la quantité en qualité ‘’ La formule laisse entendre que le changement se produit par grands sauts suivant une lente accumulation de tensions auquel un système résiste jusqu’au moment où il atteint le point de rupture. (...) Le modèle ponctué peut refléter les rythmes du changement biologique (...) ne serait-ce qu’à cause du nombre et de l’importance des résistances au changement dans les systèmes complexes à l’état stable. (...) « L’histoire de n’importe quelle région de la terre est comme la vie d’un soldat. Elle consiste en de longues périodes d’ennui entrecoupées de courtes périodes d’effroi. »
Stephen Jay Gould dans « Le pouce du panda »
Lire encore :
Le lien entre évolution non-linéaire de la Terre et évolution non-linéaire de la vie