
Accueil > 02 - Livre Deux : SCIENCES > Evolution ou révolution des espèces ? > Cinq doigts ! Pourquoi cinq ? Pour les membres antérieurs et postérieurs (...)
Cinq doigts ! Pourquoi cinq ? Pour les membres antérieurs et postérieurs de nombreux animaux et de l’homme !
mercredi 2 octobre 2019, par


Cinq doigts ! Pourquoi cinq ? Pour les membres antérieurs et postérieurs de nombreux animaux et de l’homme !
Comment se fait-il que les animaux aient souvent cinq doigts aux pieds comme aux mains, tout comme l’homme, nous demande notre lecteur J-F.P ?
Ainsi, si nous avons cinq doigts comme de nombreux animaux, est-ce parce que ce chiffre de cinq est un avantage pour la vie ? Ne serait-ce pas plutôt le fait que nous avons un ancêtre commun qui en avait cinq, c’est-à-dire du fait d’un événement de l’histoire qui n’avait pas une probabilité plus grande qu’une autre de se produire et qui n’a rien d’adaptatif ? Où est-ce que notre génétique du développement a marqué ce chiffre cinq et pourquoi ?
Beaucoup de personnes estiment que la remarque de l’apparente permanence de ces cinq doigts dans toute la lignée qui a mené à l’homme semble indiquer un sens à l’évolution, pour ne pas dire une véritable directivité, comme si l’évolution suivait un fil conducteur menant à l’homme, comme élément ultime d’une évolution dirigée ! Cette conception, contrairement aux apparences, n’est pas issue de l’observation scientifique mais des a priori idéologiques, religieux et anthropocentriques, qui atteignent non seulement le grand public mais aussi les philosophes et même les scientifiques…
Cet a priori nous dit que, si on retrouve sans cesse le même nombre de doigts, à partir d’un ancêtre commun, cette fixité signifie qu’est programmée un certain nombre d’éléments qui devront être conservés par la suite, ce qui sous-entend une certaine planification préalable du changement et du non-changement menant à un résultat final que l’on peut imaginer préétabli. Cela semble à certains comme un élément de justification des conceptions créationnistes, magiques ou mystiques. Et cela d’autant plus qu’on nous dit que les scientifiques eux-mêmes ignoreraient d’où vient ce nombre cinq qui serait systématiquement reconnu à partir d’un certain ancêtre. Où ce cinq est-il inscrit ? Dans le fonctionnement des espèces en question, dans le capital génétique, dans une nécessité physique particulière, dans un avantage évolutif retenu par la sélection ? La magie du nombre cinq peut également donner des arguments à ceux qui pensent que l’évolution cesse d’agir quand elle a atteint la perfection, argument adaptationniste (attention, l’adaptation n’est qu’une des conceptions se revendiquant du darwinisme et non la seule, ni la plus valable, comme le rappelle souvent Stephen Jay Gould) qui suggère que l’être humain détient plusieurs éléments de perfection dont les cinq doigts seraient l’un d’entre eux. Le caractère ouvertement anthropocentrique de cet argument prête à sourire évidemment… Les scientifiques disant ne pas le savoir, la place est grande pour les interprétations les plus métaphysiques. En répétant la question « pourquoi cinq », question à laquelle les études biochimiques, génétiques, physiques, embryologiques, homéogénétiques, et évolutives darwiniennes ne répondaient pas, ou semblaient ne pas répondre, ils faisaient de ce nombre un fait magique, qui justifiait un caractère supranaturel au sein même de l’évolution des espèces, une vraie justification de la prétendue « magie de la vie ».
Stephen Jay Gould dans « Comme les huit doigts de la main » :
« Darwin a soutenu que les vertébrés possèdent un plan d’organisation commun parce qu’ils descendent d’un ancêtre commun. Si les os présentent des formes et des positions semblables, c’est parce qu’ils descendent d’un ancêtre commun. Si les os présentent des formes et des positions semblables, c’est parce qu’une morphologie ancestrale est apparue de façon fortuite dans l’histoire, puis a été transmise héréditairement chez toutes les espèces faisant partie de la lignée. Cela n’a rien à voir avec la perfection éthérée d’une forme idéale d’inspiration divine…
Prenez, par exemple, le cas de ce sous-ensemble constitué par les vertébrés terrestres, un groupe techniquement appelé les Tétrapodes (= « quatre pattes ») qui comprend les amphibiens, les reptiles, les oiseaux et les mammifères, dans les classifications courantes. Certains volent, certains nagent et d’autres rampent. Par leur apparence extérieure et leur mode de vie, une baleine et un oiseau-mouche semblent suffisamment disparates pour justifier qu’on les sépare fondamentalement. Cependant, nous les réunissons dans une même catégorie classificatoire, sur la base des caractéristiques squelettiques communes à tous les tétrapodes. Celles-ci sont les traits par lesquels nous définissons aujourd’hui l’archétype de cet ensemble de vertébrés particuliers. L’un des plus importants de ces traits est la possession de quatre pattes, chacune d’elle étant munie de cinq doigts – c’est ce que l’on appelle le membre pentadactyle…
Bien qu’un oiseau-mouche ne possède que trois doigts aux pattes, l’étude de son développement embryologique permet de dire qu’il s’agit des doigts numéro deux, trois et quatre de la gamme ancestrale complète. Les traits de l’archétype fournissent des bases de départ et des configurations génératrices ; il n’est pas besoin qu’ils figurent intégralement en fin de développement.
Dans l’archétype du tétrapode, aucun trait n’a jamais paru plus canonique que le membre pentadactyle, grâce auquel, semble-t-il, tant d’activités humaines, importantes ou futiles, peuvent être menées à bien : jouer du piano, frapper sur un clavier de machine à écrire, tirer les canards à coups de fusils, acclamer en claquant des mains, compter grâce à la numération décimale… Cependant, dans ce chapitre, je vais mettre en question la façon traditionnelle d’envisager ce nombre canonique, sans nier l’importance qu’il revêt dans notre vie.
Le grand paléontologiste suédois, Erik Jarvik, a terminé son chef-d’œuvre en deux volumes sur la structure et l’évolution des vertébrés par des considérations extrêmement importantes au sujet des membres pentadactyles et des capacités humaines. Il a noté que de nombreux mammifères « évolués » ont apporté des modifications à la configuration originelle par le biais de la perte ou de la spécialisation de certains de leurs doigts – les chevaux n’en ont retenu qu’un seul, coiffé d’un sabot ; les baleines ont pratiquement perdu la totalité du membre arrière. Il a aussi remarqué que la conjugaison d’une main dotée de nombreux doigts, permettant la manipulation d’outils, et d’un cerveau de plus en plus gros, convenant bien à la découverte des meilleures façons de se servir de ces derniers et à l’invention de nouvelles techniques, s’est révélée fondamentale dans l’évolution humaine.
Si l’ancêtre de notre lignée avait perdu la flexibilité originelle du membre pentadactyle « primitif » et acquis par évolution quelque spécialisation associée à une réduction du nombre des doigts, l’intelligence humaine ne se serait jamais développée. Dans ce sens, qui est très important, nous sommes ici parce que nos ancêtres ont retenu la gamme complète archétypale de cinq doigts et ne lui ont pas substitué quelque configuration plus moderne, mais permettant finalement moins de possibilités. (…)
Est-il vrai que le membre à cinq doigts soit un trait archétypal constant et universel chez les tétrapodes, et que l’on peut l’interpréter, à la lumière de la théorie de l’évolution darwinienne, comme une configuration ancestrale qui aurait été retenue dans tous les lignages descendant des poissons ostéolépiformes ? (…)
Les nageoires des poissons sont construites sur des principes différents ; mais les ancêtres à nageoires charnues des tétrapodes ont été dotés d’une architecture osseuse qui a pu se transformer facilement pour donner les pattes avant et arrière des vertébrés terrestres. Cependant, aucun poisson n’a jamais présenté la configuration pentadactyle, et cette dernière caractéristique canonique n’est vraiment apparue qu’avec les tétrapodes.
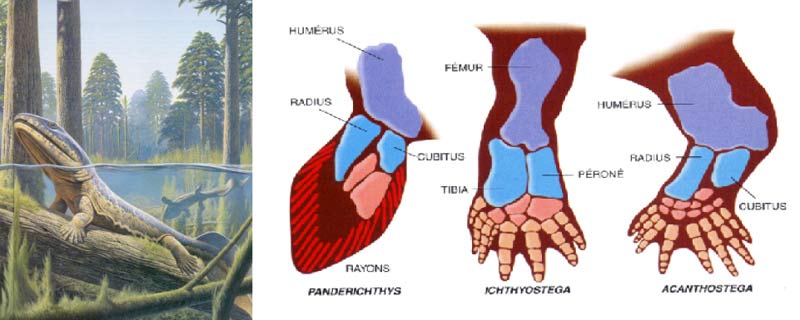
Les plus anciens tétrapodes ont été découverts dans l’Est du Groenland par une expédition danoise en 1929. Ils datent de la toute dernière phase de la période dévonienne, laquelle, s’étendant de – 390 à – 340 millions d’années, est souvent dénommée, dans les livres et les expositions de muséums, « âge des poissons » (d’après ce sot nombrilisme consistant à donner des noms aux périodes, en fonction du vertébré qui y a été, par hasard, le plus répandu).
Le paléontologiste suédois Gunnar Säve-Söderbergh a recueilli de plus abondants restes de ces animaux en 1931 et a dirigé les recherches dans cette région du Groenland jusqu’à sa mort prématurée en 1948. Erik Jarvik a alors pris sa succession et, durant les années 1950, publié ses vastes études anatomiques sur les deux genres qui se partagent le titre de tétrapode le plus ancien – Ichthyostega et Acanthostega. Bien qu’aucun des spécimens dont il disposait n’ait eu les doigts suffisamment bien conservés aux pattes avant ou arrière pour assurer un dénombrement sans ambiguïté, Jarvik a dessiné la reconstitution de ces tout premiers tétrapodes en les munissant du canonique nombre de cinq doigts par membre.
Notre confiance dans cette hypothèse non prouvée sur l’existence de cinq doigts originels a commencé à s’effriter en 1984, lorsque le paléontologiste soviétique O.A. Lebedev a rapporté que le tétrapode archaïque Tulerpeton, qui venait d’être découvert, et qui datait lui aussi de la fin du Dévonien, possédait six doigts à chacun de ses membres. Cela a conduit l’anatomiste et embryologiste J.R. Hinchliffe à suggérer en 1989 – et c’était prophétique comme nous venons juste de le vérifier – que le nombre de cinq doigts avait découlé d’un processus de stabilisation secondaire, et ne représentait pas une configuration originelle. Hinchliffe avait intitulé son article « La reconstitution de l’archétype : l’évolution du membre pentadactyle », et l’avait clos par ces mots : « In se pourrait que la réduction à la formule pentadactyle ait fait suite à une phase d’expérimentation évolutive. »
Les soupçons de Hinchliffe ont maintenant été confirmés – et au centuple. En octobre 1990, M.I. Coates et J.A. Clack ont publié leurs observations sur de nouveaux restes d’Ichthyostega et d’Acanthostega, recueillis par une expédition menée conjointement par l’université de Cambridge et une équipe de Copenhague, dans l’Est du Groenland en 1987. Quelques nouveaux spécimens remarquables – une patte arrière complète d’Ichtyostega et une patte avant d’Acanthostega – ont permis, pour la première fois, un dénombrement direct des doigts… En fait, les pattes arrière d’Ichthyostega portent, écoutez bien, sept doigts ! – trois petits doigts serrés les uns contre les autres occupant la place de l’hallux (« gros orteil », transposé en termes humains) dans la configuration à cinq doigts ordinaire aux tétrapodes. Acanthostega se démarque encore plus fortement du modèle supposé commun à tous ; sa patte antérieure comporte huit doigts, disposés selon un large arc où leur taille va d’abord croissant, puis e décroissant.
La conclusion semble inévitable et il faut carrément abandonner une vieille « certitude ». On ne connaît que trois tétrapodes dévoniens. Aucun n’est doté de membres à cinq doigts. Ils présentent, respectivement, six, sept et huit doigts, sur ceux de leurs membres que l’on a retrouvés. La configuration à cinq doigts n’est nullement canonique et archétypale pour les tétrapodes – au moins au sens « présente dès le début » (des tétrapodes). Dans le meilleur des cas, pour les fans de la pentadactylie, le nombre de cinq est issu d’un processus de stabilisation ultérieur, et ne présente pas une condition originelle.
En outre, à la lumière de ces nouvelles données, la reconsidération d’un fait connu depuis longtemps jette un doute supplémentaire sur le caractère « initial » du nombre cinq. Selon la naïve conception d’une « échelle des êtres organisés », l’évolution des vertébrés est représentée par la séquence linéaire ascendante : amphibiens-reptiles-mammifères-homme (les oiseaux constituant la seule branche latérale admise). Mais la notion d’échelle correspond à une fiction culturellement rassurante ; en réalité, l’évolution repose sur un abondant buissonnement de branches. Bien sûr, les tétrapode ont un ancêtre commun, mais les amphibiens actuels (grenouilles et salamandres) représentent le terminus d’une grande branche, et ne sont, et ne sont nullement situés à la base d’une séquence évolutive. En outre, aucun amphibien fossile ne semble être clairement l’ancêtre de la lignée des vertébrés pleinement terrestres (reptiles, oiseaux et mammifères) appelés amniotes en l’honneur de l’œuf « amniote » (enveloppé d’une coquille dure et possédant sa « mare interne »), innovation évolutive qui a permis, selon les métaphores couramment en usage, « la conquête complète du milieu terrestre » ou « la vraie libération par rapport à l’eau ».
Il y a là une question annexe par rapport au sujet traité dans cet article, mais arrêtez-vous un instant et réfléchissez aux préjugés intrinsèques à de telles « descriptions ». Pourquoi la possibilité de pondre des œufs sur la terre ferme est-elle une « libération » ; est-ce que l’eau équivaut à un esclavage ? Pourquoi le fait de n’habiter exclusivement que dans un seul type de milieu est-il une « conquête » ? Qui se bat pour quoi ? De telles formulations n’ont de sens que si l’évolution des êtres vivants s’efforce de « monter » vers l’apogée humaine – conception de l’évolution la plus sotte et anthropocentrique que je puisse imaginer.
Les premiers reptiles fossiles sont à peu près aussi vieux que les premiers amphibiens situés clairement dans la lignée qui a finalement donné nos grenouilles et salamandre actuelles. Ainsi, plutôt qu’une échelle menant des amphibiens aux reptiles, les archives fossiles, tout comme l’étude des vertébrés actuels, suggèrent que le tronc des tétrapodes s’est scindé très tôt en deux branches principales – les amphibiens et les amniotes (reptiles, oiseaux et mammifères).
Venons-en maintenant à la question de la pentadactylie et de ses limites : les amniotes présentent effectivement le modèle canonique de cinq doigts à chaque membre (avec parfois des modifications superposées à cet état initial). Mais les amphibiens, à la fois vivants et fossiles, possèdent cinq doigts à leurs pattes arrière et seulement QUATRE à leurs pattes avant. Les anatomistes savent cela depuis des années, mais ont toujours supposé que cette réduction à quatre avait succédé à un stade initial canonique de cinq. Il faut maintenant remettre en question cette hypothèse. Si tous les premiers tétrapodes possédaient plus de cinq doigts, et si les premiers amphibiens ont été séparés des amniotes dès le début de la vie terrestre, pourquoi vouloir que la configuration à quatre doigts des pattes avant des amphibiens soit descendue d’une formule initiale de cinq ? Le nombre de cinq chez tous les tétrapodes actuels est probablement issu d’un processus de stabilisation à partir d’un nombre plus élevé. Il se pourrait que la formule de quatre doigts aux pattes antérieures chez les amphibiens soit également directement issue de ce nombre supérieur, sans qu’il n’y ait jamais eu de stade pentadactyle entre les deux. S’il en est ainsi, la pentadactylie est une notion qui ne tient pas pour deux raisons :
1) elle ne représente pas l’état originel des tétrapodes (ainsi que le prouve l’existence des plus anciennes formes à six, sept ou huit doigts.
2) elle ne correspond pas à une formule canonique chez l’une des deux grandes lignées des tétrapodes.
Un brillant papier sur le développement embryologique des membres a fourni des éléments permettant de bien comprendre ces nouvelles conceptions. Cet article a rapporté des travaux réalisés par Neil H. Shbin et Père Albrech. Shubin et Albrech ont essayé de rendre compte de la complexité du membre des tétrapodes, en expliquant qu’il résultait de l’interaction de trois processus fondamentaux : un processus de division dichotomique (une série d’éléments donnant deux séries) ; un processus de segmentation (un élément donnant d’autres éléments au sein d’une série) ; et un processus de condensation (plusieurs éléments fusionnant entre eux). Le membre se construit en direction de l’extérieur du corps – de l’épaule jusqu’aux doigts de la main ; de la cuisse jusqu’aux orteils. Le processus commence par l’édification d’un élément unique partant du tronc – l’humérus pour le bras ; le fémur pour la jambe. Un phénomène de division dichotomique engendre les éléments suivants dans la séquence – le radius et le cubitus pour le bras ; le tibia et le péroné pour la jambe. Le processus de division dichotomique (jusqu’aux os du poignet) est le phénomène fondamental qui conduit à la formation des doigts. Il est très nettement asymétrique, dans la mesure où l’un des os cesse de se diviser (pour ne donner qu’une seule rangée de segments, tandis que le membre continue à se développer), alors que l’autre est à l’origine de toutes les autres multiplications d’éléments suivantes, y compris la production de doigts. Assez étrangement, l’os qui ne donne lieu à aucune division est le plus gros des deux éléments – le radius, dans le cas du bras ; et le tibia, dans celui de la jambe. La main et le pied sont issus d’un processus de divisions dichotomiques procédant à partir du plus petit élément – le cubitus dans le cas du bras ; et le péroné dans celui de la jambe…
Dans le cadre de la conception traditionnelle, on pouvait parler d’un doigt dominant (support de l’axe central) et d’éléments subordonnés (les éléments qui étaient issus de divisions dichotomiques se réalisant à des distances de plus en plus grandes), mais on ne pouvait pas rapporter ce modèle à quelque programmation temporelle que ce soit. Dans le cadre du modèle révisé par Shubin et Albrech, la série des doigts répond à une séquence temporelle : la position spatiale est en rapport avec l’ordre temporel de la formation. Il y a une équivalence entre « en arrière » et « ancien » ; « en avant » et « récent »…
Le modèle de Shubin et Albrech suggère un mécanisme simple et évident pour expliquer la stabilisation du nombre de doigts à cinq après une période initiale de labilité, où ce nombre pouvait être plus élevé et varié. Si les doigts se forment de l’arrière vers l’avant selon une séquence temporelle, alors on peut facilement arriver à une réduction du nombre des doigts par un arrêt plus précoce de la séquence. Ce principe est évident et très général : arrêtez plus tôt ! (…) L’évolution peut réduire le nombre de doigts en arrêtant le mécanisme générateur lorsqu’il a atteint le nombre de cinq dans sa marche d’arrière en avant…
Cette manière de voir permet de rendre compte immédiatement de certaines vieilles données de l’histoire naturelle, restées jusque-là inexpliquées. Dans tous les groupes de tétrapodes, de nombreuses lignées ont réduit la formule originelle de cinq doigts à un nombre plus petit – quelquefois jusqu’au minimum de un, comme chez le cheval… Sur la base du modèle de Shubin et d’Albrech, la raison de cette règle est évidente : dernier formé, premier éliminé… Le phénomène inverse des mutations polydactyles (donnant plus que cinq doigts) va aussi dans le sens du modèle de Shubin et Albrech. Chez l’homme, la plupart des mutations polydactyliques engendrent un sixième doigt en tant que simple duplication de l’un d’eux au sein de la gamme habituelle de cinq. Et, puisque cet événement se produit après la série des divisions dichotomiques donnant initialement le chiffre de cinq, il n’entre pas dans le cadre des phénomènes envisagés par Shubin et Albrech. Mais, dans plusieurs autres espèces, les doigts surnuméraires sont engendrés par le déroulement de la séquence génératrice au-delà de la formation du doigt numéro un, qui marque généralement la fin de la série.
J.R. Hincliffe a écrit en 1989 : « De nombreux mutants polydactyles… possèdent la série des cinq doigts normaux et y ajoutent des doigts surnuméraires en position préaxiale (c’est-à-dire que ces derniers sont formés après l’élaboration du doigt numéro un). » En outre, Hincliffe cite quelques données expérimentales sur l’inhibition de la synthèse de l’ADN pendant le développement embryologique du lézard Lacerta viridis. Dans la mesure où il y a moins de matériel disponible pour l’édification des organes, des doigts peuvent être alors perdus. Le doigt formé en dernier est toujours le premier à s’en aller.
Les données provenant de deux types d’observations différentes soutiennent l’idée que les doigts sont formés par le biais d’un processus générateur séquentiel, opérant d’arrière en avant, et que la position d’un doigt dans l’espace est en corrélation avec sa position dans le déroulement temporel de l’édification embryologique : les doigts supplémentaires sont ajoutés, et les doigts anciens perdus, à partir du doigt qui représente le point terminal de la séquence normale – le doigt numéro un. (…)
Nous avions l’habitude de penser que la formule de cinq doigts était invariante et canonique, et notre principale question était toujours : pourquoi cinq ? Mais si ce dernier nombre résulte d’un processus de stabilisation secondaire, d’un point d’arrêt dans le déroulement d’une séquence temporelle qui pourrait aller plus loin (mais ne le fait pas), il faut se poser une question très différente et de bien des façons plus intéressante : pourquoi s’arrêter en ce point ? (…)
Alors, pourquoi cinq doigts ? Il existe deux grands types de réponses à cette question. L’une, darwinienne classique, ou adaptationniste, essaie d’apercevoir un avantage marqué, ou même une inévitabilité, à la formule des cinq doigts, en termes d’utilité dans le cadre du mode de vie d’un animal (avantage qui pourrait peut-être promouvoir cette configuration par le biais de la sélection naturelle). On peut présenter des arguments plausibles sur les bénéfices qu’elle pourrait apporter dans le cadre de la vie sur les continents. Les êtres vivants que l’évolution a fait passer de l’eau sur la terre émergée ont dû affronter de nombreux problèmes inédits, dont aucun n’était plus sévère que la force de gravité, nouvelle pour eux, et ont eu à soutenir le poids de leur corps en l’absence de la force ascensionnelle procurée antérieurement par l’eau. Le remplacement des nageoires par des membres a fourni le moyen d’assurer ce soutien, et, selon un argument avancé depuis longtemps, il se pourrait que la configuration de cinq éléments ait été optimale pour supporter le poids du corps – car comprenant un axe central représenté par le doigt numéro trois, et deux contreforts puissants et symétriques de chaque côté… Sur la base de cette argumentation, les tétrapodes possèderaient cinq doigts, parce que le soutien du corps et la locomotion imposeraient (ou du moins encourageraient fortement) cette formule en tant que solution optimale…
Le contre-argument le plus évident peut aussi, paradoxalement, venir soutenir l’adaptationnisme : si la configuration de cinq doigts est optimale sur la terre ferme, pourquoi les tout premiers tétrapodes possédaient respectivement six, sept et huit doigts à leurs pattes ? Une réponse adaptationniste consiste à dire que ces animaux ont acquis leurs membres dans le cadre de la locomotion dans l’eau car ils menaient, de façon prédominante, voire totalement, un mode de vie aquatique. Et, en effet, comme cela a été reconnu depuis longtemps, de sérieux indices suggèrent qu’Ichtyostega vivait en grande partie dans l’eau : il présentait une petite nageoire caudale et un système de « ligne latérale » sur le crâne… La reconstitution des membres d’Ichtyostega et d’Acanthostega effectuée par Coates et Clack va également dans le sens de l’hypothèse du mode de vie aquatique avancée ci-dessus : la forme de ces organes était hydrodynamique et leur capacité de rotation était limitée, de sorte qu’ils restaient en position horizontale, à la manière des nageoires, et n’étaient pas dirigés vers le bas de façon à soutenir le poids du corps au-dessus du sol…
Mais de nombreux éléments peuvent aussi mettre fortement en doute cette explication adaptationniste. Premièrement, comme nous l’avons dit ci-dessus, certains membres de la lignée tétrapode, les amphibiens, ne possèdent que quatre doigts à leurs pattes avant, et il n’y a pas de preuves, dans ce cas, d’une phase originelle à cinq doigts. Donc la pentadactylie n’a peut-être pas été un stade universel chez les vertébrés terrestres. Deuxièmement, si la configuration de cinq doigts confère un avantage dans le soutien du poids du corps, pourquoi notre espèce préférée, servant de base de référence traditionnelle à toute chose – Homo sapiens – l’a-t-elle conservée au niveau de tous ses membres, alors que son organisation anatomique ne fait appel qu’aux deux membres postérieurs pour contrer les forces de gravité ? Et d’ailleurs, les cinq éléments à l’extrémité de ces derniers ne sont pas organisés symétriquement autour d’un puissant doigt central, car c’est le gros orteil qui est le principal soutien du poids corporel.
Le deuxième grand type de réponse à la question : « pourquoi cinq doigts ? » repose sur ce que j’ai appelé la contingence historique (voir mon livre « La vie est belle »). Elle consiste à dire que la configuration de cinq doigts n’a pas été expressément façonnée, mais s’est simplement trouvée là à un moment donné. D’autres configurations auraient pu fonctionner et auraient peut-être pu être édifiées par l’évolution, mais il n’en a rien été – et cinq doigts convenaient suffisamment bien…
Si la configuration de cinq doigts est si bonne que cela, pourquoi de nombreuses espèces recourent à des moyens vraiment curieux et retors pour arriver à produire six doigts (grâce à un prepollux ou à un os du poignet reconverti) ? Si elle est prévisible à ce point, pourquoi l’une des deux lignées de tétrapodes n’a-t-elle développé que quatre doigts aux pattes avant ? (…)
Mais l’argument qui va le plus dans le sens de la contingence est celui qui a déterminé l’écriture de cet essai – la découverte des sept doigts chez Ichthyostega et de huit chez Acanthyostega. Si les tétrapodes avaient eu cinq doigts dès le début, et avaient toujours conservé cinq doigts par la suite, alors, on pourrait légitimement invoquer une certaine prévisibilité ou inévitabilité… Mais si les premiers membres de la lignée avaient six, sept ou huit doigts, alors, des quantités d’autres possibilités étaient envisageables, et la configuration de cinq a sans doute été retenue fortuitement non par nécessité…
La notion de contingence est riche et fascinante : elle renvoie à une subtile interaction entre la capacité des individus à modifier l’histoire et les limites bien définies posées par les lois de la nature… Les événements contingents ont forgé notre monde à tous et la nature. »
Stephen Jay Gould, « Le pouce du panda » :
« Les pandas passent le plus clair de leur temps à dévorer leur cher bambou. Assis bien droit sur leur derrière, ils manipulent leurs tiges avec leurs pattes avant, se débarrassant des feuilles pour ne consommer que les pousses. (…) Comment le descendant d’une lignée adaptée à la course peut utiliser ses mains de façon si habile ? Ils tiennent les tiges de bambou dans leurs pattes et les dépouillent de leurs feuilles en faisant passer les tiges entre un pouce apparemment flexible et les autres doigts. (…) L’adroite utilisation d’un pouce opposable compte parmi les marques du génie humain. Nous avons maintenu, exagéré même, cette importante flexibilité de nos ancêtres primates, alors que la plupart des mammifères l’ont sacrifiée en spécialisant leurs doigts. Les carnivores courent, griffent et grattent. Mon chat peut me manipuler psychologiquement, mais jamais il ne tapera à la machine ni ne jouera du piano. Aussi ai-je compté les autres doigts du panda pour m’apercevoir – O surprise plus grande encore ! - qu’ils étaient au nombre de cinq et non de quatre. Ce pouce était-il un sixième doigt qui aurait évolué séparément ?
(…) Anatomiquement, le « pouce » du panda n’est pas un doigt. Il est construit à partir d’un os appelé le sésamoïde radial (du radius), normalement un des petits os formant le poignet. Chez le panda, le sésamoïde radial est très développé et si allongé que sa taille atteint presque celle des os des phalanges des vrais doigts. Le sésamoïde radial soutient un renflement de la patte avant du panda (…) Le pouce du panda est doté non seulement d’un os pour lui assurer sa force, mais également de muscles pour assurer son agilité. Ces muscles, comme le sésamoïde radial lui-même, n’ont pas été créés de toutes pièces. Comme les organes des orchidées de Darwin, ce sont des éléments anatomiques communs, remodelés pour une fonction nouvelle. (…) L’anatomie des autres carnivores nous fournit-elle une indication sur l’origine de cette curieuse disposition chez les pandas ? (…) Chez la plupart des carnivores, ces mêmes muscles qui, chez le panda, agissent sur le sésamoïde radial, sont attachés uniquement à la base du pollex, ou vrai pouce. Mais, chez les ours communs, le long abducteur se termine par deux tendons : l’un s’insère à la base du pouce, comme chez la plupart des carnivores, mais l’autre est fixé au sésamoïde radial. (…) Il semble que toute la succession des transformations de la musculature ait découlé automatiquement d’une simple hypertrophie de l’os sésamoïde. Le pouce sésamoïde du panda est une structure complexe formée par le développement prononcé d’un os et par une profonde redisposition de la musculature. (…) L’allongement du sésamoïde radial a pu être provoqué par une transformation génétique, peut-être une seule mutation affectant le rythme et la vitesse de la croissance. (…) Le vrai pouce du panda, trop spécialisé pour être utilisé à une autre fonction et devenir un doigt opposable, apte à la manipulation, est relégué à un autre rôle. Le panda est donc contraint de se servir des organes disponibles et de choisir cet os du poignet hypertrophié, solution quelque peu bâtarde mais très fonctionnelle. Le pouce du panda ne remportera pas de prix au concours Lépine de la nature. Selon l’expression de Michael Ghiselin, ce n’est qu’un truc et non un mécanisme élégant. Mais il atteint le but recherché et nous passionne d’autant plus que ses éléments de départ ne sont pas ceux que l’on aurait pu imaginer. Le traité de Darwin sur les orchidées est rempli d’illustrations similaires. Le souci Epipactis, par exemple, se sert de son labelle – un pétale agrandi – comme d’un piège. (…) L’insecte doit sortir par une seule issue qui lui est offerte, ce qui le force à se frotter contre les masses de pollen. (…) Darwin montre comment, chez des orchidées, le même labelle évolue pour entrer dans la composition d’une série de systèmes ingénieux dont le but est d’assurer la fécondation croisée.(…) Toutes ces adaptations ont eu comme point de départ un organe qui n’était autre, chez quelque lointaine forme ancestrale, qu’un pétale conventionnel. (…) Darwin écrit : « Bien qu’un organe ait pu, à l’origine, ne pas être formé dans un but bien précis, s’il remplit à présent cette fonction, nous pouvons dire, à juste titre, qu’il a été spécialement conçu pour cela. Selon le même principe, si un homme a fabriqué une machine dans un but bien précis, mais a utilisé pour sa construction de vieilles roues et poulies, des ressorts usagers, en ne leur faisant subir que de légères modifications, on doit dire de cette machine dans son ensemble, avec toutes ses pièces constitutives, qu’elle a été spécialement conçue dans le but visé. Ainsi, dans la nature tout entière, presque tous les organes de chaque être vivant ont probablement servi, dans des conditions légèrement modifiées, à des buts divers, et ont joué un rôle dans la machinerie vivante de nombreuses formes spécifiques anciennes, distinctes des formes actuelles. » Sans doute la métaphore des roues et des poulies rafistolées n’est-elle guère flatteuse mais nous devons surtout porter attention au résultat obtenu. La nature, selon le mot de François Jacob, est un excellent bricoleur et non un artisan divin. Et qui peut se permettre de mettre en doute le bon fonctionnement de ces quelques cas exemplaires ? »
Que dit la génétique ?
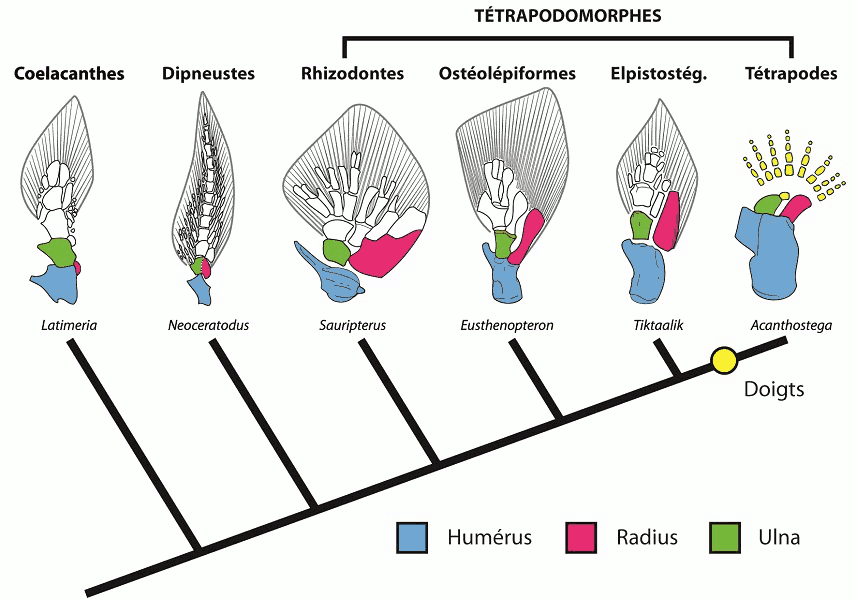
La pentadactylie des êtres humains est souvent considérée comme un caractère ancestral plésiomorphique issu du membre chiridien des tétrapodes du Dévonien (370 ma), membre dérivant originairement de la « nageoire » aux nombreux rayons (nageoire pectorale ou ventrale) des anciens poissons marins sarcoptérygiens.
Les membres des fossiles de tétrapodes aquatiques présentent une polydactylie à 8, 7, 6 ou doigts (Ichthyostega qui possède sept orteils et Tulerpeton six doigts) qui implique le gène HOXD13.
L’explication de la pentadactylie actuelle en termes d’évolution est qu’elle est probablement issue d’un processus de stabilisation à partir d’un nombre de doigts plus grand, faisant intervenir le gène hoxa11.
Plus particulièrement, deux gènes avait déjà été identifiés, les gènes hoxa13 et hoxd13, comme étant responsables de la formation des nageoires et des membres. "On sait que chez la souris, si les gènes hoxa13 et hoxd13 sont désactivés, on obtient des animaux qui n’ont pas de doigts. Et il y a un autre travail qui est paru dans Nature il y a quelques semaines qui a montré que si on désactive ces deux mêmes gènes chez les poissons, alors leurs nageoires n’ont plus de rayons" explique Marie Kmita, auteure principale de l’étude publiée dans Nature.
Sauf que ces gènes, s’ils sont une preuve du lien entre nageoires et membres, n’expliquent pas pourquoi nous avons cinq doigts et non pas sept ou huit. D’ailleurs, comme le souligne le site Science Daily, des fossiles remontant à plusieurs millions d’années montrent que parmi les premières espèces sur terre, certaines possédaient plus de six doigts. Alors comment expliquer que nous n’en avons que cinq ?
Marie Kmita, directrice de l’IRCM, et Yacine Kherdjemil, l’un de ses doctorants, ont génétiquement modifié des souris. Ils ont découvert qu’une troisième version des gènes "hox", le hoxa11, avait un rôle primordial dans le nombre de doigts que peuvent posséder les vertébrés.
C’est en essayant de reproduire les mêmes versions de gènes hox des poissons qu’ils ont fait ce constat. Chez les poissons notamment, les gènes hoxa11 et hoxa13 sont actifs dans des cellules des nageoires en développement. Alors que chez les souris, ils sont actifs dans d’autres cellules que celles des membres. Or, lorsque les chercheurs ont activé le gène hoxa11 dans des cellules allant devenir des doigts, ils ont obtenu des souris avec six ou sept doigts par membre. "Quand il est actif dans les cellules qui vont devenir les doigts, on obtient plus de 5 doigts, et quand on le désactive, les pattes des souris redeviennent normales", détaille Marie Kmita à La Presse.
"Cela suggère que ce changement morphologique majeur n’a pas eu lieu par l’acquisition de nouveaux gènes mais seulement par la modification de leurs activités", explique-t-elle aussi à Science Daily. Cette découverte pourrait aider la médecine à comprendre pourquoi certaines malformations génitales durant la grossesse ne sont pas uniquement le résultat de mutations mais aussi d’expression et de contrôle des gènes.
En août dernier, des chercheurs de Chicago ont démontré que deux gènes – Hoxa13 et Hoxd13 – sont responsables de la formation des rayons des nageoires et de nos doigts. « Ce résultat est très excitant, car il établit clairement un lien moléculaire entre les rayons des nageoires et les doigts », dit Yacine Kherdjemil, étudiant au doctorat dans le laboratoire de Marie Kmita et premier auteur de l’article paru dans Nature.
Cependant, la transition de la nageoire au membre ne s’est pas faite d’un coup. L’analyse des fossiles nous indique que nos ancêtres étaient polydactyles, c’est-à-dire qu’ils avaient un nombre de doigts supérieurs à cinq, et soulève donc une autre question clé. Par quel mécanisme l’évolution a-t-elle favorisé la pentadactylie (cinq doigts) chez les espèces actuelles ?
Une observation a particulièrement retenu l’attention de l’équipe de la Dre Kmita : « Au cours du développement, chez la souris comme chez l’homme, les gènes Hoxa11 et Hoxa13 sont activés dans des domaines séparés du bourgeon de membre, alors que, chez le poisson, ces gènes sont activés dans des domaines chevauchant la nageoire en développement », mentionne Marie Kmita, directrice de l’unité de recherche en génétique et développement de l’Institut de recherches cliniques de Montréal (IRCM) et professeure-chercheuse à la Faculté de médecine de l’Université de Montréal.
En essayant de comprendre la signification de cette différence, Yacine Kherdjemil a démontré que, en reproduisant l’activité de type poisson (chevauchant) pour le gène Hoxa11, les souris développent jusqu’à sept doigts par patte, c’est-à-dire un retour au statut ancestral. L’équipe de la Dre Marie Kmita a également découvert la séquence d’ADN responsable de la transition de l’activité du gène Hoxa11 de type poisson et souris. « Cela signifie que ce changement morphologique majeur ne s’est sans doute pas fait par l’acquisition de nouveaux gènes, mais simplement par la modification de leurs activités », ajoute la Dre Marie Kmita.
Quelques remarques supplémentaires
Les tétrapodes actuels descendent d’un ancêtre commun pentadactyle (nous ne descendons donc pas d’Acanthostega, qui avait huit doigts ou Ichthyostega qui en avait sept, le plus ancien fossile adapté à la locomotion terrestre et avec une pentadactylie fonctionnelle étant Pederpes), et, au cours de l’évolution, il a été plus facile d’introduire des variations sur le même thème que de reconstruire de novo des membres pour chaque usage : le membre pentadactyle a été "facilement" modifié durant l’évolution, s’adaptant par sélections successives aux besoins liés au mode de vie de l’espèce. Ainsi, la pentadactylie des mammifères n’est probablement que la « nécessité impérative d’une organisation précise de l’appareil urogénital, de notre tractus digestif ou de notre colonne vertébrale, puisque les mêmes gènes officient dans toutes ces structures. Il est donc vraisemblablement vain de disserter sur la valeur adaptative de la pentadactylie ».
L’ancêtre reptilien direct des dinosaures théropodes est un petit carnivore bipède qui marche comme un oiseau. (Les théropodes sont un des groupes de vertébrés les plus florissants et le clade de dinosaures le plus diversifié morphologiquement)
• Ses pattes avant sont libres pour saisir des objets ou des proies, et le plus long doigt est le deuxième, non le troisième comme chez les autres Sauropsides. Il a toutefois cinq doigts par main, alors que presque tous les théropodes primitifs et les oiseaux en ont trois. Chez cet animal, l’articulation de la cheville est déjà en charnière, et les métatarses sont allongés, relevés : l’animal marche, comme les dinosaures après lui. Les nombreuses modifications ultérieures du pied permettent probablement une foulée plus longue et plus rapide, qualité qui aidera ultérieurement les théropodes à développer le vol.
• Les premiers théropodes présentent des adaptations qui allègent leur squelette : des os creux et des cavités dans le crâne.
• Comme les oiseaux actuels, leur cou est long et leur dos horizontal. Le quatrième doigt de la main des premiers dinosaures est déjà réduit, et le cinquième doigt a déjà presque disparu. Le quatrième doigt devient bientôt lui-aussi un moignon, puis ces deux doigts réduits disparaissent ensemble chez les théropodes tétanoures. Les trois doigts restants fusionnent au cours de l’évolution des oiseaux postérieurs à Archaeopteryx.
• Les membres postérieurs des premiers théropodes se transforment aussi. Ils s’allongent : le fémur devient plus long que le tibia, et la fibula se réduit.
• Ces dinosaures marchent sur les trois doigts du milieu, ceux qu’utilisent toujours les oiseaux actuels. Le cinquième orteil, sans articulation, est raccourci et effilé. Le premier orteil, qui dépasse sur le côté du deuxième orteil, est formé d’un os raccourci, d’une articulation et d’une griffe. Il est placé au-dessus des autres orteils, tel un ergot.






Messages
1. Cinq doigts ! Pourquoi cinq ? Pour les membres antérieurs et postérieurs de nombreux animaux et de l’homme !, 2 octobre 2019, 06:57, par JFP/Jean-François POULIQUEN.
JFP¦¦021092019¦¦Cinq doigts à chaque membre peut être un vrai questionnement, mais l’interrogation initiale et la relation de départ de votre lecteur J-F.P était la base décimale et les dix doigts de nos deux mains. Bel article ‼‼
▬Amicalement. Les jumeaux JFP/Jean-François POULIQUEN
1. Cinq doigts ! Pourquoi cinq ? Pour les membres antérieurs et postérieurs de nombreux animaux et de l’homme !, 2 octobre 2019, 08:02, par Robert Paris
Certes, certes, mais la réponse est là : raisonnerions-nous en base dix si nous n’avions pas deux fois cinq doigts aux mains ?
2. Cinq doigts ! Pourquoi cinq ? Pour les membres antérieurs et postérieurs de nombreux animaux et de l’homme !, 2 octobre 2019, 19:08, par JFP/Jean-François POULIQUEN.
JFP¦¦021092019¦¦Si nous avions un nombre de doigts différent de 5 doigts à chaque main, et bien non, nous ne raisonnerions pas en base 10, car si 1,2, 3 ou même 4 doigts et bien Ɂ mais si 6 doigts par main, et bien la base 12 serait sûrement utilisée je pense, et après on se limiterai forcément, car déjà 12 est important et on inventerai pas plus de 12 signes ‼‼ Sur un nuage de 10 choses nous ne savons pas évaluer d’un coup d’œil le nombre de ces 10 choses, car il faut une gymnastique d’esprit et de concentration pour pouvoir voir les 10 choses, et plus c’est forcément non, et il en va aussi pour 8 et donc 4 doigts, mais que sans doute que cette base de 8 serait possible et aurait été retenue, car on peut la diviser par 2 puis encore par 2, comme en binaire... Dans tous les cas un groupement de 4 et plus de choses, nous ne savons pas évaluer d’un coup d’œil le nombre de ces choses, mais nous c’est nous les humains, car certains animaux savent repérer des groupes d’entités importantes,, bien plus que nous. Si on perçoit d’un seul coup d’œil une deux ou trois choses, à partit de quatre choses cela se complique, et pourtant quatre c’est très peu. Pourquoi et bien nous n’en savons rien, d’autant que des animaux savent repérer d’un coup d’œil plus de choses que nous ‼‼
▬Amicalement. Les jumeaux JFP/Jean-François POULIQUEN